5.3 Bacterial Transformation
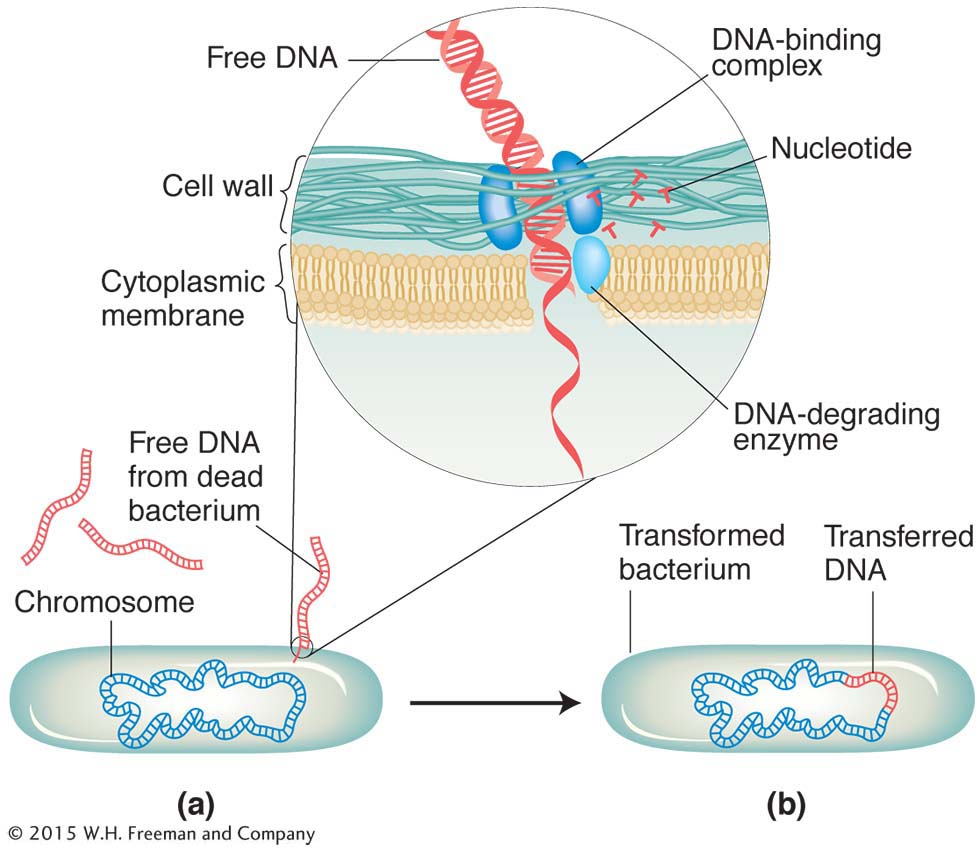
Some bacteria can take up fragments of DNA from the external medium, and such uptake constitutes another way in which bacteria can exchange their genes. The source of the DNA can be other cells of the same species or cells of other species. In some cases, the DNA has been released from dead cells; in other cases, the DNA has been secreted from live bacterial cells. The DNA taken up integrates into the recipient’s chromosome. If this DNA is of a different genotype from that of the recipient, the genotype of the recipient can become permanently changed, a process aptly termed transformation.
The nature of transformation
Transformation was discovered in the bacterium Streptococcus pneumoniae in 1928 by Frederick Griffith. Later, in 1944, Oswald T. Avery, Colin M. MacLeod, and Maclyn McCarty demonstrated that the “transforming principle” was DNA. Both results are milestones in the elucidation of the molecular nature of genes. We consider this work in more detail in Chapter 7.
The transforming DNA is incorporated into the bacterial chromosome by a process analogous to the double-
Transformation has been a handy tool in several areas of bacterial research because the genotype of a strain can be deliberately changed in a very specific way by transforming with an appropriate DNA fragment. For example, transformation is used widely in genetic engineering. It has been found that even eukaryotic cells can be transformed, by using quite similar procedures, and this technique has been invaluable for modifying eukaryotic cells (see Chapter 10).
Chromosome mapping using transformation
Transformation can be used to measure how closely two genes are linked on a bacterial chromosome. When DNA (the bacterial chromosome) is extracted for transformation experiments, some breakage into smaller pieces is inevitable. If two donor genes are located close together on the chromosome, there is a good chance that sometimes they will be carried on the same piece of transforming DNA. Hence, both will be taken up, causing a double transformation. Conversely, if genes are widely separated on the chromosome, they will most likely be carried on separate transforming segments. A genome could possibly take up both segments independently, creating a double transformant, but that outcome is not likely. Hence, in widely separated genes, the frequency of double transformants will equal the product of the single-
Unfortunately, the situation is made more complex by several factors—