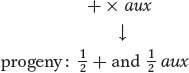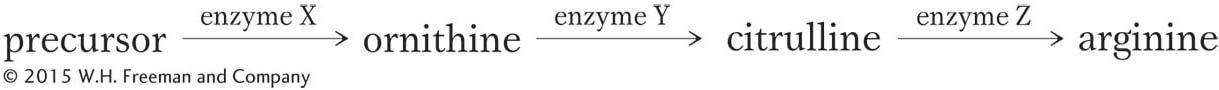6.2 Interaction of Genes in Pathways

Figure 6-9: Discoverer of inborn errors of metabolism
Figure 6-9: British physician Archibald Garrod (1857-1936).
[Science Photo Library/Science Source.]
Genes act by controlling cellular chemistry. Early in the twentieth century, Archibald Garrod, an English physician (Figure 6-9), made the first observation supporting this insight. Garrod noted that several recessive human diseases show defects in what is called metabolism, the general set of chemical reactions taking place in an organism. This observation led to the notion that such genetic diseases are “inborn errors of metabolism.” Garrod worked on a disease called alkaptonuria (AKU), or black urine disease. He discovered that the substance responsible for black urine was homogentisic acid, which is present in high amounts and secreted into the urine in AKU patients. He knew that, in unaffected people, homogentisic acid is converted into maleylacetoacetic acid; so he proposed that, in AKU, there is a defect in this conversion. Consequently, homogentisic acid builds up and is excreted. Garrod’s observations raised the possibility that the cell’s chemical pathways were under the control of a large set of interacting genes. However, the direct demonstration of this control was provided by the later work of Beadle and Tatum on the fungus Neurospora.
Biosynthetic pathways in Neurospora
The landmark study by George Beadle and Edward Tatum in the 1940s not only clarified the role of genes, but also demonstrated the interaction of genes in biochemical pathways. They later received a Nobel Prize for their study, which marks the beginning of all molecular biology. Beadle and Tatum did their work on the haploid fungus Neurospora, which we have met in earlier chapters. Their plan was to investigate the genetic control of cellular chemistry. In what has become the standard forward genetic approach, they first irradiated Neurospora cells to produce mutations and then tested cultures grown from ascospores for interesting mutant phenotypes relevant to biochemical function. They found numerous mutants that had defective nutrition. Specifically, these mutants were auxotrophic mutants, of the type described for bacteria in Chapter 5. Whereas wild-type Neurospora can use its cellular biochemistry to synthesize virtually all its cellular components from the inorganic nutrients and a carbon source in the medium, auxotrophic mutants cannot. In order to grow, such mutants require a nutrient to be supplied (a nutrient that a wild-type fungus is able to synthesize for itself), suggesting that the mutant is defective for some normal synthetic step.
As their first step, Beadle and Tatum confirmed that each mutation that generated a nutrient requirement was inherited as a single-gene mutation because each gave a 1:1 ratio when crossed with a wild type. Letting aux represent an auxotrophic mutation,
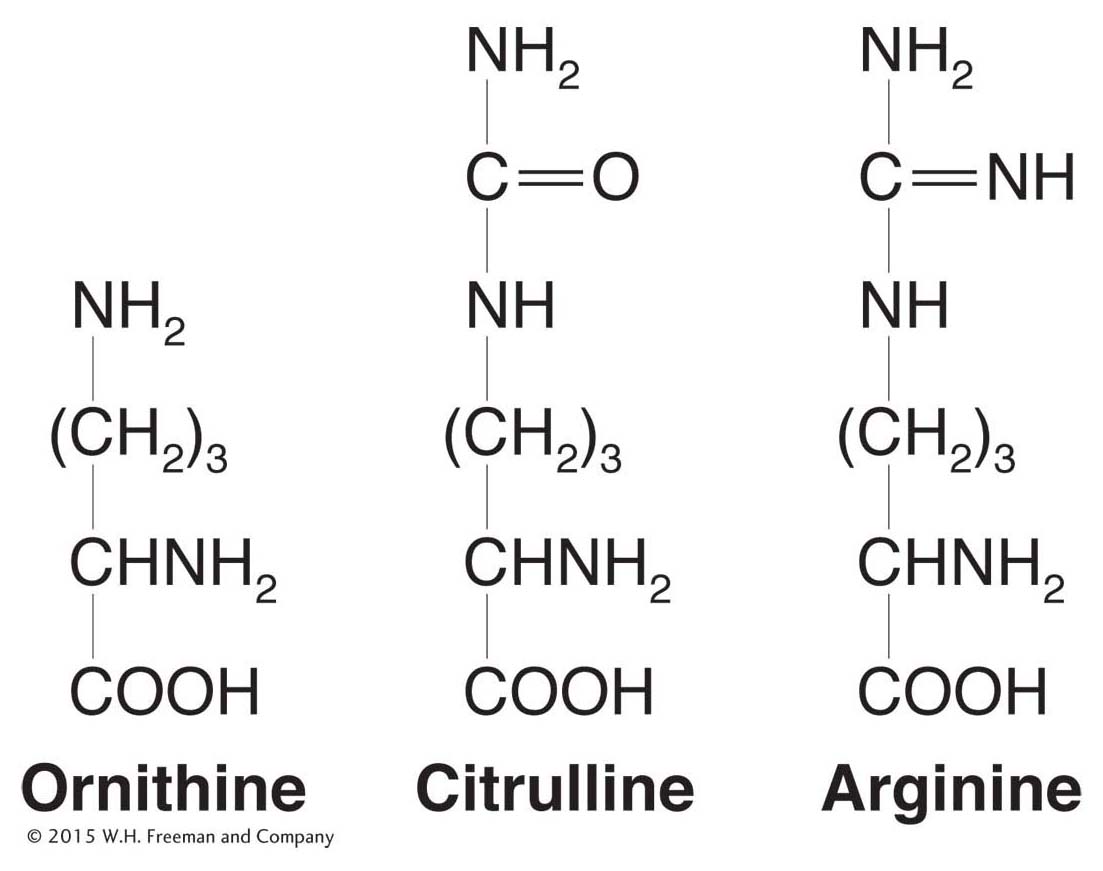
Figure 6-10: Arginine and its analogs
Figure 6-10: The chemical structures of arginine and the structurally related compounds citrulline and ornithine.
Their second step was to classify the specific nutritional requirement of each auxotroph. Some would grow only if proline was supplied, others methionine, others pyridoxine, others arginine, and so on. Beadle and Tatum decided to focus on arginine auxotrophs. They found that the genes that mutated to give arginine auxotrophs mapped to three different loci on three separate chromosomes. Let’s call the genes at the three loci the arg-1, arg-2, and arg-3 genes. A key breakthrough was Beadle and Tatum’s discovery that the auxotrophs for each of the three loci differed in their response to the structurally related compounds ornithine and citrulline (Figure 6-10). The arg-1 mutants grew when supplied with any one of the chemicals ornithine, citrulline, or arginine. The arg-2 mutants grew when given arginine or citrulline but not ornithine. The arg-3 mutants grew only when arginine was supplied. These results are summarized in Table 6-1.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Note: A plus sign means growth; a minus sign means no growth. |
Table 6-1: Growth of arg Mutuants in Response to supplements
Cellular enzymes were already known to interconvert such related compounds. On the basis of the properties of the arg mutants, Beadle and Tatum and their colleagues proposed a biochemical pathway for such conversions in Neurospora:
This pathway nicely explains the three classes of mutants shown in Table 6-1. Under the model, the arg-1 mutants have a defective enzyme X, and so they are unable to convert the precursor into ornithine as the first step in producing arginine. However, they have normal enzymes Y and Z, and so the arg-1 mutants are able to produce arginine if supplied with either ornithine or citrulline. Similarly, the arg-2 mutants lack enzyme Y, and the arg-3 mutants lack enzyme Z. Thus, a mutation at a particular gene is assumed to interfere with the production of a single enzyme. The defective enzyme creates a block in some biosynthetic pathway. The block can be circumvented by supplying to the cells any compound that normally comes after the block in the pathway.
We can now diagram a more complete biochemical model:
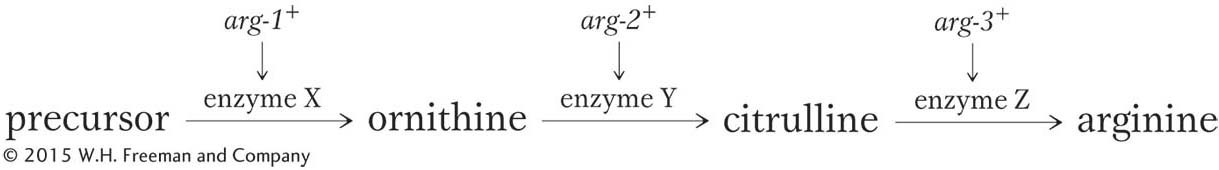
This brilliant model, which was initially known as the one-gene-one-enzyme hypothesis, was the source of the first exciting insight into the functions of genes: genes somehow were responsible for the function of enzymes, and each gene apparently controlled one specific enzyme in a series of interconnected steps in a biochemical pathway. Other researchers obtained similar results for other biosynthetic pathways, and the hypothesis soon achieved general acceptance. All proteins, whether or not they are enzymes, also were found to be encoded by genes, and so the phrase was refined to become the one-gene-one-polypeptide hypothesis. (Recall that a polypeptide is the simplest type of protein, a single chain of amino acids.) It soon became clear that a gene encodes the physical structure of a protein, which in turn dictates its function. Beadle and Tatum’s hypothesis became one of the great unifying concepts in biology because it provided a bridge that brought together the two major research areas of genetics and biochemistry.
We must add parenthetically that, although the great majority of genes encode proteins, some are now known to encode RNAs that have special functions. All genes are transcribed to make RNA. Protein-encoding genes are transcribed to messenger RNA (mRNA), which is then translated into protein. However, the RNA encoded by a minority of genes is never translated into protein because the RNA itself has a unique function. These are called functional RNAs. Some examples are transfer RNAs, ribosomal RNAs, and small cytoplasmic RNAs—more about them in later chapters.
KEY CONCEPT
Chemical synthesis in cells is by pathways of sequential steps catalyzed by enzymes. The genes encoding the enzymes of a specific pathway constitute a functionally interacting subset of the genome.
Gene interaction in other types of pathways
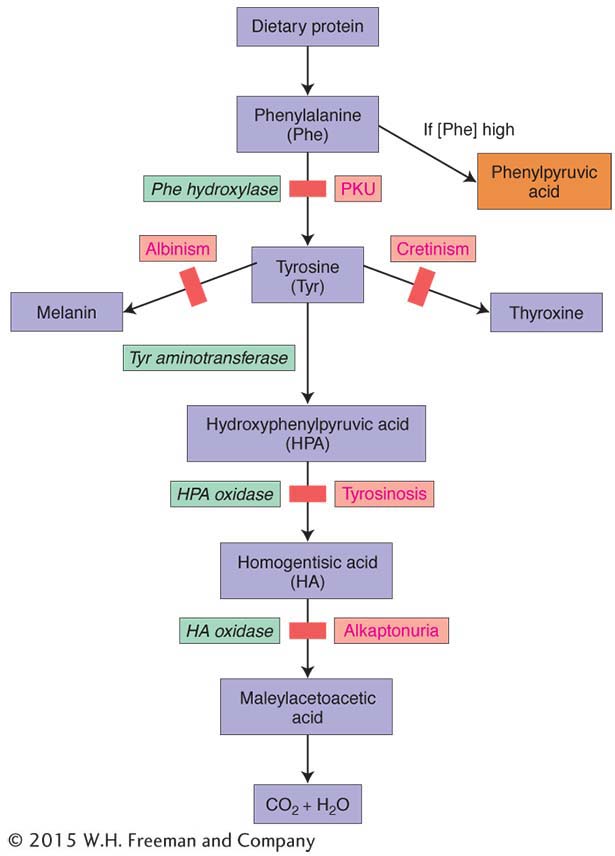
Figure 6-11: A synthetic pathway and associated diseases
Figure 6-11: A section of the phenylalanine metabolic pathway in humans, including diseases associated with enzyme blockages. The disease PKU is produced when the enzyme phenylalanine hydroxylase malfunctions. Accumulation of phenylalanine results in an increase in phenylpyruvic acid, which interferes with the development of the nervous system.
The notion that genes interact through pathways is a powerful one that finds application in all organisms. The Neurospora arginine pathway is an example of a synthetic pathway, a chain of enzymatic conversions that synthesizes essential molecules. We can extend the idea again to a human case already introduced, the disease phenylketonuria (PKU), which is caused by an autosomal recessive allele. This disease results from an inability to convert phenylalanine into tyrosine. As a result of the block, phenylalanine accumulates and is spontaneously converted into a toxic compound, phenylpyruvic acid. The PKU gene is part of a metabolic pathway like the Neurospora arginine pathway, and part of it is shown in Figure 6-11. The illustration includes several other diseases caused by blockages in steps in this pathway (including alkaptonuria, the disease investigated by Garrod).
Another type of pathway is a signal-transduction pathway. This type of pathway is a chain of complex signals from the environment to the genome and from one gene to another. These pathways are crucial to the proper function of an organism. One of the best understood signal-transduction pathways was worked out from a genetic analysis of the mating response in baker’s yeast. Two mating types, determined by the alleles MATa and MATα, are necessary for yeast mating to occur. When a cell is in the presence of another cell of opposite mating type, it undergoes a series of changes in shape and behavior preparatory to mating. The mating response is triggered by a signal-transduction pathway requiring the sequential action of a set of genes. This set of genes was discovered through a standard interaction analysis of mutants with aberrant mating response (most were sterile). The steps were pieced together by using the approaches in the next section. The signal that sets things in motion is a mating pheromone (hormone) released by the opposite mating type; the pheromone binds to a membrane receptor, which is coupled to a G protein inside the membrane and activates the protein. The G protein, in turn, sets in motion a series of sequential protein phosphorylations called a kinase cascade. Ultimately, the cascade activates the transcription of a set of mating-specific genes that enable the cell to mate. A mutation at any one of these steps may disrupt the mating process.
Developmental pathways comprise the steps by which a zygote becomes an adult organism. This process involves many genetically controlled steps, including establishment of the anterior-posterior and dorsal-ventral axes, laying down the basic body plan of organs, and tissue differentiation and movement. These steps can require gene regulation and signal transduction. Developmental pathways will be taken up in detail in Chapter 13, but the interaction of genes in these pathways is analyzed in the same way, as we will see next.