Chapter Introduction
The Genetic Control of Development
The Genetic Control
of Development
LEARNING OUTCOMES
After completing this chapter, you will be able to
Distinguish members of the genetic toolkit for development from other genes and explain how they are identified.
Correlate where and when pattern-regulating genes are expressed during development with the phenotypes that result from mutations in them.
Explain examples of how spatially restricted patterns of gene expression are generated during development.
Relate the biochemical functions of toolkit proteins to their effects on the development of bodies or body parts.
Identify homologous components of the genetic toolkit in different animal phyla.
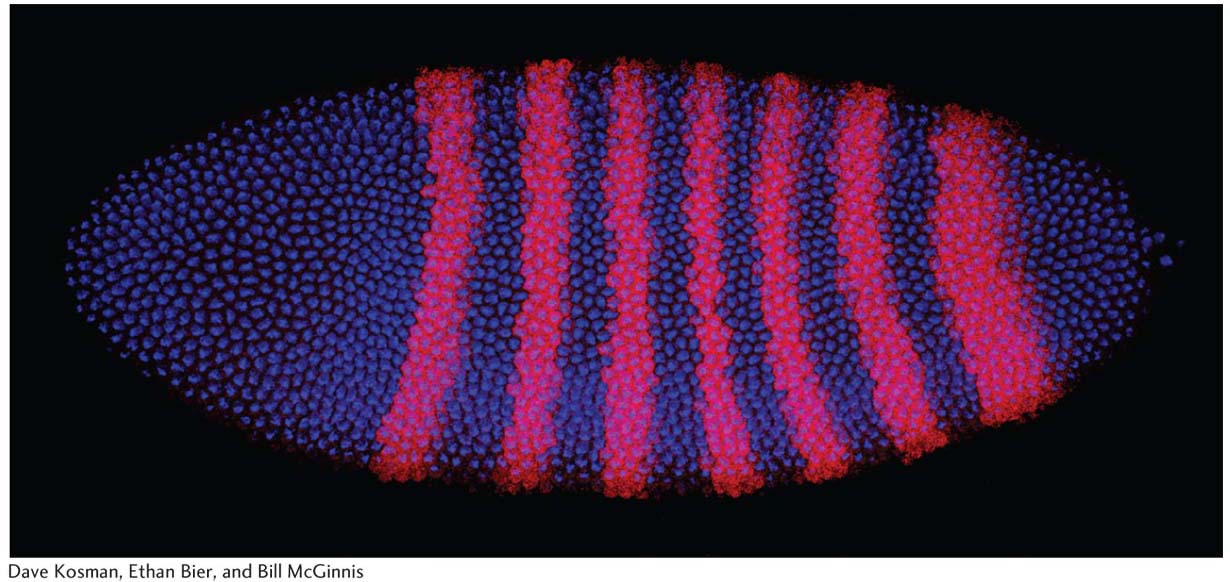
Gene expression in a developing fruit-fly embryo. The seven magenta stripes mark the cells expressing the mRNA of a gene encoding a regulatory protein that controls segment number in the Drosophila embryo. The spatial regulation of gene expression is central to the control of animal development.
[Dave Kosman, Ethan Bier, and Bill McGinnis.]
OUTLINE
|
|
The genetic approach to development |
|
|
The genetic toolkit for Drosophila development |
|
|
Defining the entire toolkit |
|
|
Spatial regulation of gene expression in development |
|
|
Post-transcriptional regulation of gene expression in development |
|
|
From flies to fingers, feathers, and floor plates: the many roles of individual toolkit genes |
|
|
|
Of all the phenomena in biology, few if any inspire more awe than the formation of a complex animal from a single-celled egg. In this spectacular transformation, unseen forces organize the dividing mass of cells into a form with a distinct head and tail, various appendages, and many organs. The great geneticist Thomas Hunt Morgan was not immune to its aesthetic appeal:

Figure 13-1: Homeotic mutants of Drosophila melanogaster
Figure 13-1: In homeotic mutants, the identity of one body structure has been changed into another. (a) Normal fly with one pair of forewings on the second thoracic segment and one pair of small hind wings on the third thoracic segment. (b) Triple mutant for three mutations in the Ultrabithorax gene. Ubx function is lost in the posterior thorax, which causes the development of forewings in place of the hind wings. (c) Antennapedia mutant in which the antennae are transformed into legs.
[Sean Carroll.]
A transparent egg as it develops is one of the most fascinating objects in the world of living beings. The continuous change in form that takes place from hour to hour puzzles us by its very simplicity. The geometric patterns that present themselves at every turn invite mathematical analysis…. This pageant makes an irresistible appeal to the emotional and artistic sides of our nature.1
Yet, for all its beauty and fascination, biologists were stumped for many decades concerning how biological form is generated during development. Morgan also said that “if the mystery that surrounds embryology is ever to come within our comprehension, we must …have recourse to other means than description of the passing show.”
The long drought in embryology lasted well beyond Morgan’s heyday in the 1910s and 1920s, but it was eventually broken by geneticists working very much in the tradition of Morgan-style genetics and with his favorite, most productive genetic model, the fruit fly Drosophila melanogaster.
The key catalysts to understanding the making of animal forms were the discoveries of genetic monsters—mutant fruit flies with dramatic alterations of body structures (Figure 13-1). In the early days of Drosophila genetics, rare mutants arose spontaneously or as by-products of other experiments with spectacular transformations of body parts. In 1915, Calvin Bridges, then Morgan’s student, isolated a fly having a mutation that caused the tiny hind wings (halteres) of the fruit fly to resemble the large forewings. He dubbed the mutant bithorax. The transformation in bithorax mutants is called homeotic (Greek homeos, meaning same or similar) because one part of the body (the hind wing) is transformed to resemble another (the forewing), as shown in Figure 13-1b. Subsequently, several more homeotic mutants were identified in Drosophila, such as the dramatic Antennapedia mutant in which legs develop in place of the antennae (Figure 13-1c).
The spectacular effects of homeotic mutants inspired what would become a revolution in embryology, once the tools of molecular biology became available to understand what homeotic genes encoded and how they exerted such enormous influence on the development of entire body parts. Surprisingly, these strange fruit-fly genes turned out to be a passport to the study of the entire animal kingdom, as counterparts to these genes were discovered that played similar roles in almost all animals.
The study of animal and plant development is a very large and still-growing discipline. As such, we do not attempt a comprehensive overview of embryology. Rather, in this chapter, we will focus on a few general concepts that illustrate the logic of the genetic control of animal development. We will explore how the information for building complex structures is encoded in the genome. In contrast to the control of gene regulation in single bacterial or eukaryotic cells, the genetic control of body formation and body patterning is fundamentally a matter of gene regulation in three-dimensional space and over time. Yet we will see that the principles governing the genetic control of development are connected to those already presented in Chapters 11 and 12, governing the physiological control of gene expression in bacteria and single-celled eukaryotes.