Chapter Introduction
Mutation, Repair, and Recombination
581
Mutation, Repair,
and Recombination
CHAPTER
16
LEARNING OUTCOMES
After completing this chapter, you will be able to
Explain the molecular basis of mutations.
Compare and contrast the origins and outcomes of spontaneous versus induced mutations.
Describe the different biological repair mechanisms.
Describe human genetic diseases that are caused by mutations in repair mechanisms.
Discuss the differences between cancerous and normal cells.
Explain why mutagenic agents can cause some cancers.

OUTLINE
16.1 The phenotypic consequences of DNA mutations
16.2 The molecular basis of spontaneous mutations
16.3 The molecular basis of induced mutations
16.4 Biological repair mechanisms
16.5 Cancer: an important phenotypic consequence of mutation
582
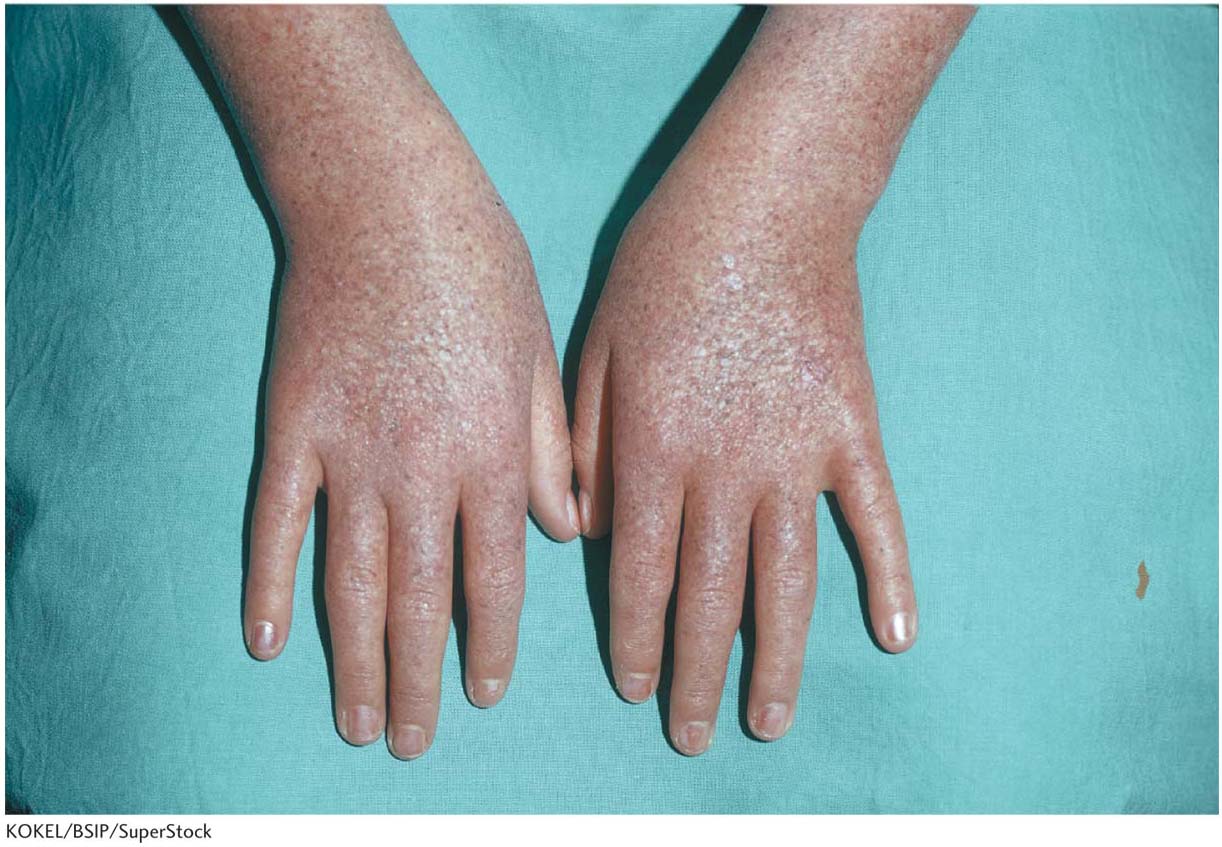
A young patient develops a great many small, frecklelike, precancerous skin growths and is extremely sensitive to sunlight (Figure 16-1). A family history is taken, and the patient is diagnosed with an autosomal recessive disease called xeroderma pigmentosum. Throughout her life, she will be prone to developing pigmented skin cancers. Several different genes can be mutated to generate this disease phenotype. In a person without the disease, each of these genes contributes to the biochemical processes in the cell that respond to chemical damage to DNA and repair this damage before it leads to the formation of new mutations. Later in this chapter, we will see how mutations in the repair systems lead to genetic diseases such as xeroderma pigmentosum.
Persons with this disease are examples of genetic variants—individuals that show phenotypic differences in one or more particular characters. Because genetics is the study of inherited differences, genetic analysis would not be possible without variants. In preceding chapters you saw many analyses of the inheritance of such variants; now, we consider their origin. How do genetic variants arise?
Two major processes are responsible for genetic variation: mutation and recombination. We have seen that mutation is a change in the DNA sequence of a gene. Mutation is especially significant because it is the ultimate source of evolutionary change; new alleles arise in all organisms, some spontaneously and others resulting from exposure to radiation or chemicals in the environment. The new alleles produced by mutation become the raw material for a second level of variation, effected by recombination. As its name suggests, recombination is the outcome of cellular processes that cause alleles of different genes to become grouped in new combinations (see Chapter 4). To use an analogy, mutation occasionally produces a new playing card, but it is recombination that shuffles the cards and deals them out as different hands.
In the cellular environment, DNA molecules are not absolutely stable: each base pair in a DNA double helix has a certain probability of mutating. As we will see, the term mutation covers a broad array of different kinds of changes. These changes range from the simple swapping of one base pair for another to the disappearance of an entire chromosome. In Chapter 17, we will consider mutational changes that affect entire chromosomes or large pieces of chromosomes. In the present chapter, we focus on mutational events that take place within individual genes. We call such events gene mutations.
Cells have evolved sophisticated systems to identify and repair damaged DNA, thereby preventing the occurrence of most but not all mutations. We can view DNA as being subjected to a dynamic tug-
We will see that the most potentially serious class of DNA damage, a double-
583