Modern organisms provide clues about the evolution of cell–cell interactions and multicellularity
Multicellular organisms can indeed be multicellular—
Migration of cells to form a group
Organization of cells in the cluster
Specialization of cells into tissues
Connections between cells
Even though single-
Studying the evolutionary origin of multicellularity is a challenge because it happened so long ago. The closest unicellular relatives of most modern animals and plants probably existed hundreds of millions of years ago. The transition from single-
Aggregation of cells into a cluster
Intercellular communication within the cluster
Specialization of some cells within the cluster
Organization of specialized cells into groups (tissues)
Does this list look familiar? (See above for the origin of organs in the embryo.)
A key event would have been the evolution of intercellular communication, which is necessary to coordinate the activities of different cells within a multicellular organism.
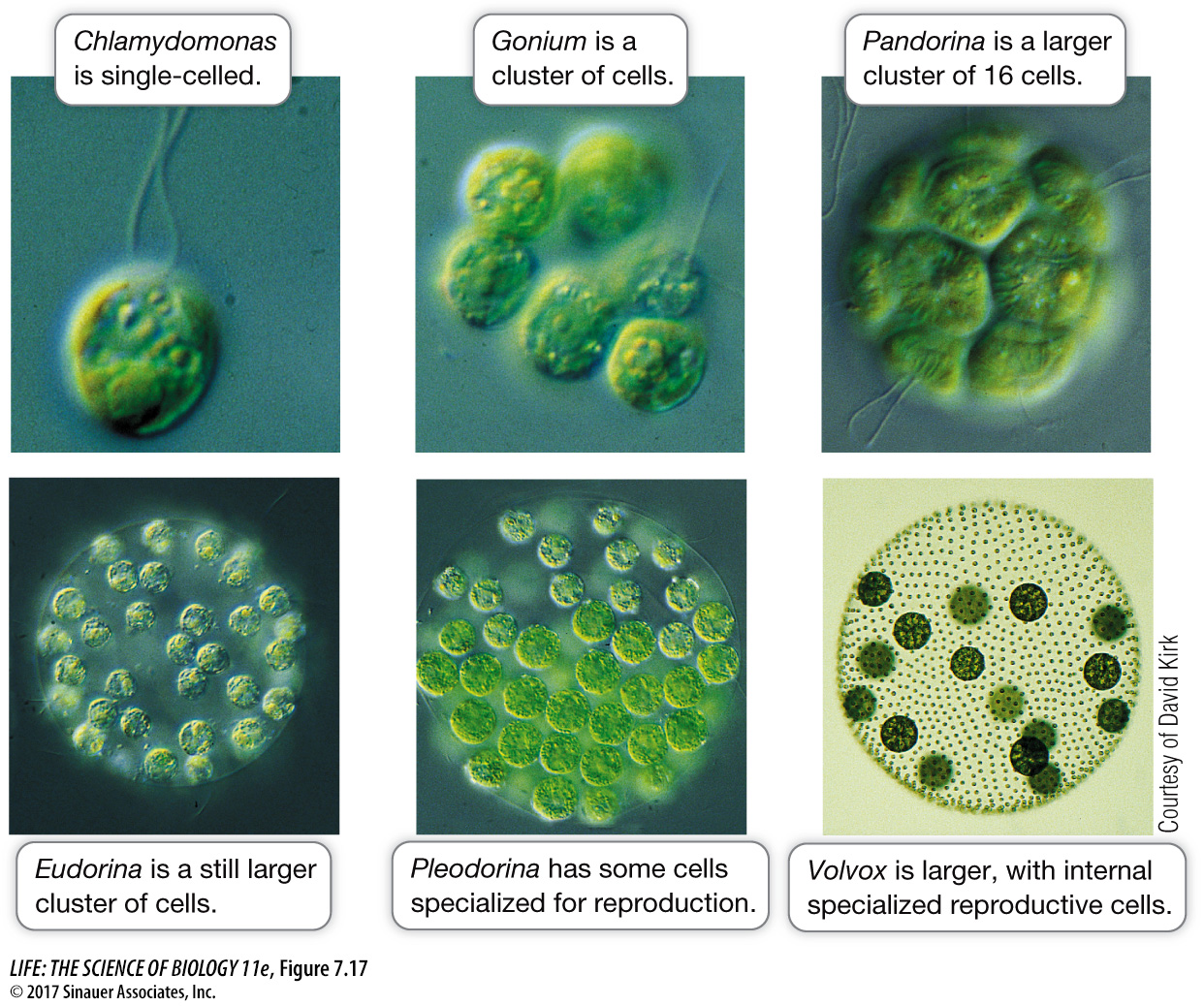
We can visualize how the evolution of multicellularity might have occurred by looking at the “Volvocine line” of aquatic green algae (Chlorophyta). These plants range from single cells to complex multicellular organisms with differentiated cell clusters (Figure 7.17). Included in this range are a single-
Q: Why was the evolution of direct communication important for tissue formation?
Direct communication between cells allows them to rapidly share signals, which travel from one cell to another in a group. This can result in common activities for a group of cells, which is important for tissues.
Chlamydomonas is the single-
Media Clip 7.1 Social Amoebas Aggregate on Cue
The separation of somatic and reproductive functions in Volvox is possible because of a key intercellular signaling mechanism that coordinates the activities of the separate tissues within the organism. Volvox has a gene whose protein product is produced by the outer, motile cells and travels to the reproductive cells, causing them to lose their flagella and divide. This gene is not active in species such as Gonium and Pandorina, which show cell aggregation but no cell specialization.