Experiments on bread mold established that genes determine enzymes
As they work to explain the principles that govern life, biologists often turn to organisms that are easy to manipulate experimentally. Such model organisms have certain characteristics that make them attractive experimental subjects. For example, model organisms:
are easy to grow in the laboratory or greenhouse.
have short generation times.
are easy to manipulate genetically, by crossing or by other methods.
often produce large numbers of progeny.
Biologists have used model organisms to develop principles of genetics that can then be applied more generally to other organisms. You have seen some of these organisms in previous chapters:
Pea plants (Pisum sativum) were used by Mendel in his genetics experiments.
Fruit flies (Drosophila) were used by Morgan in his genetics experiments.
Escherichia coli was used by Meselson and Stahl to study DNA replication.
To this list we now add the bread mold Neurospora, an ascomycete fungus (see Chapter 30). This mold is haploid for most of its life cycle, so there are no dominant or recessive alleles: all alleles are expressed phenotypically and not masked by a heterozygous condition. Neurospora is easy to grow in the laboratory. Biologists at Stanford University led by George Beadle and Edward Tatum undertook studies to biochemically define the phenotypes in Neurospora.
Like Garrod, Beadle and Tatum hypothesized that the expression of a specific gene results in the activity of a specific enzyme. They set out to test this hypothesis directly. They grew Neurospora on a nutritional medium containing sucrose, minerals, and biotin, which is the only vitamin that wild-
The scientists then treated the wild-
Mutations provide a powerful way to determine cause and effect in biology. Nowhere has this been more evident than in the elucidation of biochemical pathways. Such pathways consist of sequential events (chemical reactions) in which each event is dependent on the occurrence of the preceding event. The general reasoning is as follows:
Observation. A particular gene (a) is present and a particular reaction catalyzed by a particular enzyme (A) occurs; the two are correlated.
Hypothesis. Gene a determines the synthesis of enzyme A.
Test of hypothesis. Mutate gene a. Prediction: no functional enzyme is made, and the reaction does not occur.
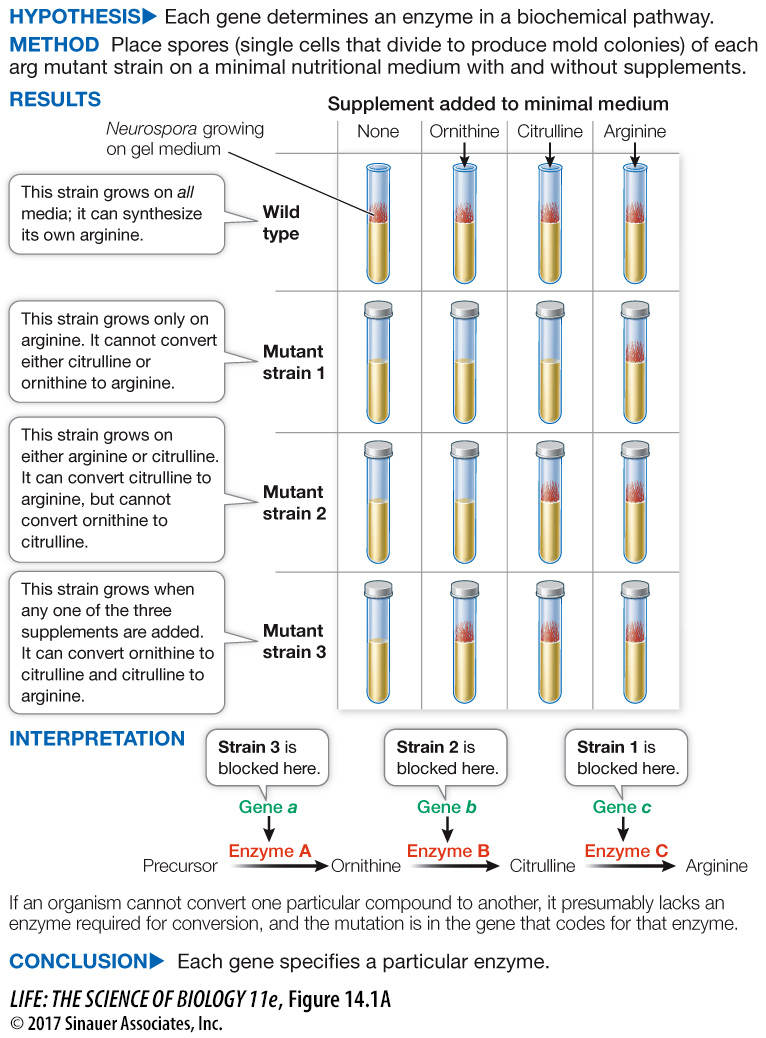
Two colleagues of Beadle and Tatum, Adrian Srb and Norman Horowitz, used this experimental approach to isolate Neurospora mutants that could not survive without the amino acid arginine in their growth medium. By adding particular compounds to the medium, Srb and Horowitz were able to identify a series of steps in the biochemical pathway leading to the synthesis of arginine (Figure 14.1a).
experiment
Figure 14.1A One Gene, One Enzyme
Original Paper: Srb, A. M. and N. H. Horowitz. 1944. The ornithine cycle in Neospora and its genetic control. Journal of Biological Chemistry 154: 129–
Srb and Horowitz developed several mutant strains of Neurospora that could not make arginine (arg). Several molecules including ornithine and citrulline are needed for arginine synthesis. By systematically adding each of these molecules to the growth media for the mutant strains, the researchers deduced that each mutant strain was deficient in one enzyme along a biochemical pathway.
work with the data
Figure 14.1B One Gene, One Enzyme
Original Paper: Srb, A. M. and N. H. Horowitz. 1944.
Neurospora (left) is haploid for most of its life cycle, except for the formation of a diploid cell when it undergoes mating; the cell then undergoes meiosis to form haploid spores. Beadle and Tatum used X rays to cause mutations in Neurospora. They isolated mutant strains that were unable to grow on minimal medium but were able to grow if the medium was supplemented with particular compounds. Their colleagues Adrian Srb and Norman Horowitz analyzed 15 mutant strains (the arg mutants) that could not synthesize arginine but could grow on medium supplemented with arginine. The scientists tested various compounds and found two, ornithine and citrulline, that could be used instead of arginine to support the growth of some of the mutant strains (as seen Figure 14.1A). The results for three of the strains are shown in the first three rows of the table, with growth expressed as dry weight of fungal material after growth for 5 days.
| Strain | No addition | Ornithine added | Citrulline added | Arginine added |
|---|---|---|---|---|
| 34105 | 1.1 | 25.5 | 30.0 | 33.2 |
| 33442 | 2.3 | 2.5 | 42.7 | 43.8 |
| 36703 | 0.0 | 0.0 | 0.0 | 20.4 |
| Double mutant | 0.0 | 0.0 | 0.0 | 22.0 |
QUESTIONS
1.
Based on the biochemical pathway for arginine synthesis shown in the experiment (Figure 14.1A), which gene (a, b, or c) encoding proteins (A, B, or C) was mutated in each strain?
34105: gene a
33442: gene b
36703: gene c
2.
Why was there some growth in strains 34105 and 33442 even when there were no additions to the growth medium?
The mutant strains may have had some residual enzyme activity, allowing for growth.
3.
Nineteen other amino acids were tested as substitutes for arginine in the three strains. In all cases, there was no growth. Explain these results.
Arginine is part of proteins and is essential for the tertiary structure of proteins. Replacing it with other amino acids is not possible.
4.
Sexual reproduction in Neurospora was used to create double mutants, which carried the mutations from both parental strains. A double mutant derived from strains 33442 and 36703 had the growth characteristics shown in the last row of the table. Explain these data in terms of the genes, mutations, and biochemical pathway.
The double mutant cannot do the reactions ornithine → citrulline and citrulline → arginine. So adding ornithine or citrulline does not allow growth, since arginine is not produced.
A similar work with the data exercise may be assigned in LaunchPad.