Regulating gene transcription conserves energy
As a normal inhabitant of the human intestine, E. coli must be able to adjust to sudden changes in its chemical environment. Its host may present it with one foodstuff one hour (e.g., glucose in fruit) and another the next (e.g., lactose in milk). Such changes in nutrients present the bacterium with a metabolic challenge. Glucose is its preferred energy source, and is the easiest sugar to metabolize. Lactose is a β-galactoside—
β-Galactoside permease is a carrier protein in the bacterial cell membrane that moves the sugar into the cell.
β-Galactosidase is an enzyme that hydrolyses lactose to glucose and galactose.
β-Galactoside transacetylase transfers acetyl groups from acetyl CoA to certain β-galactosides. Its role in the metabolism of lactose is not clear.
When E. coli grows and reproduces in a lab medium that contains glucose but no lactose or other β-galactosides, the levels of these three proteins are extremely low—
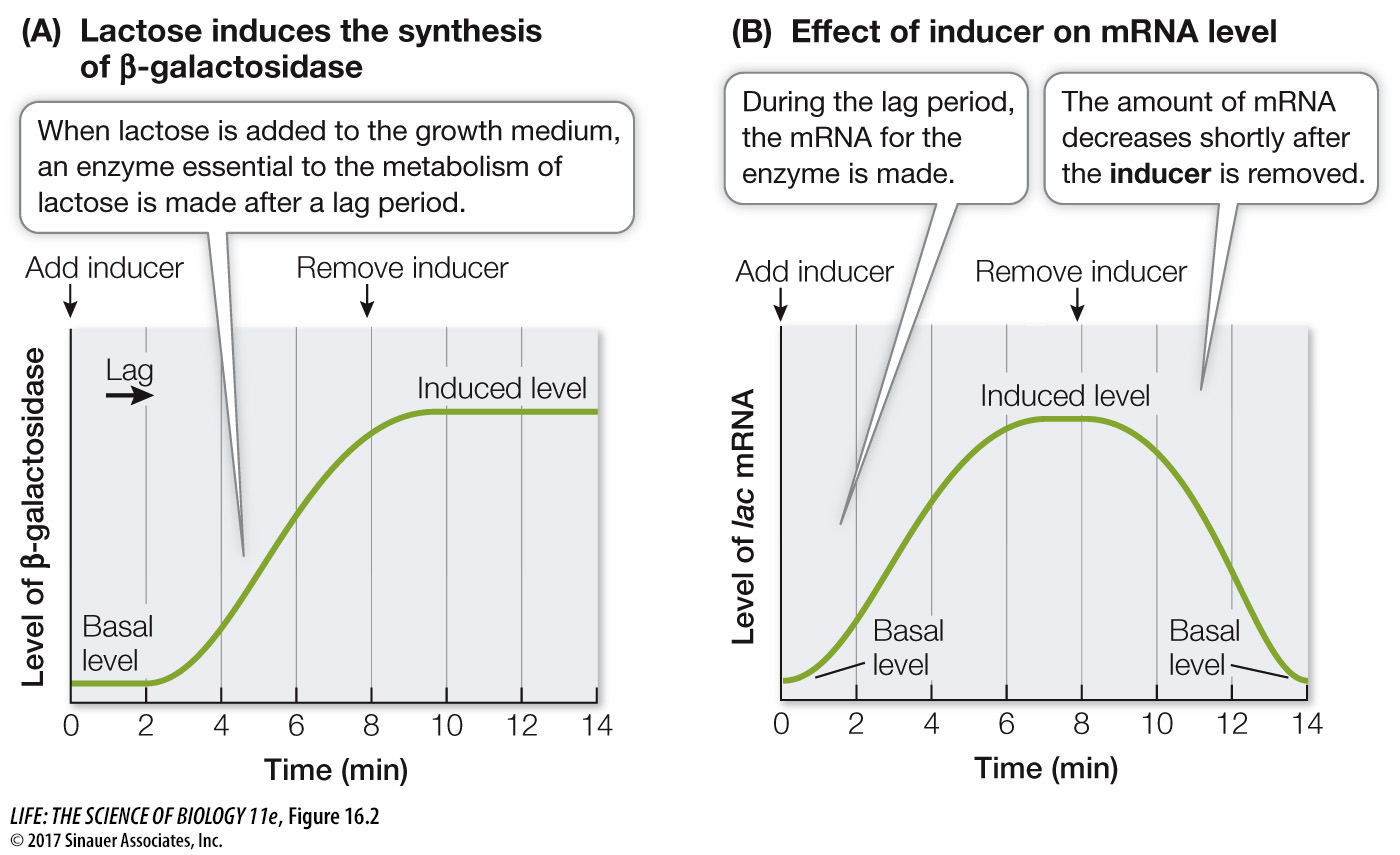
What’s behind this dramatic increase? An important clue comes from measuring the amount of mRNA for β-galactosidase. The mRNA level increases during the lag period after lactose is added to the medium, and this mRNA is translated into protein (Figure 16.2B). Moreover, the high mRNA level depends on the presence of lactose, because if the lactose is removed, the mRNA level goes down. The response of the bacterial cell to lactose is clearly at the level of transcription.
Compounds such as lactose that stimulate the synthesis of a protein are called inducers. The proteins that are produced are called inducible proteins, whereas proteins that are made all the time at a constant rate are called constitutive proteins. (Think of the constitution of a country, a document that does not change under normal circumstances.)
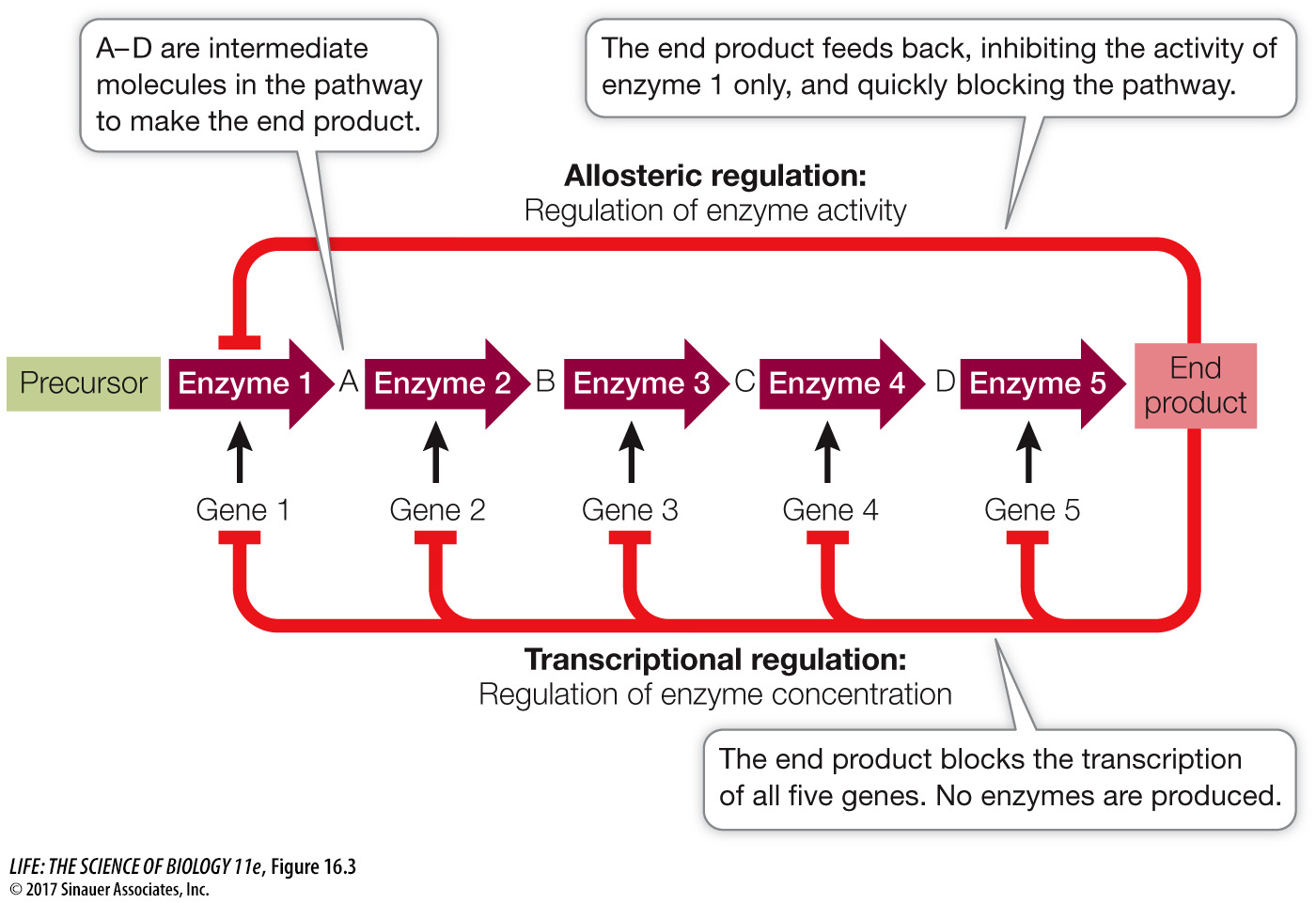
We have now seen two basic ways of regulating the rate of a metabolic pathway. In Key Concept 8.5 we described the allosteric regulation of enzyme activity, which allows the rapid fine-