Modularity allows for differences in the patterns of gene expression
The modularity of development means that the molecular pathways for developmental processes such as organ formation operate independently from one another. For instance, the Antennapedia mutant fly shown on p. 414 grew a leg where an antenna should be, but the rest of the segments formed structures normally. Modularity allows the timing and position of a developmental process to change without changing the whole organism. Several patterns have been noted.
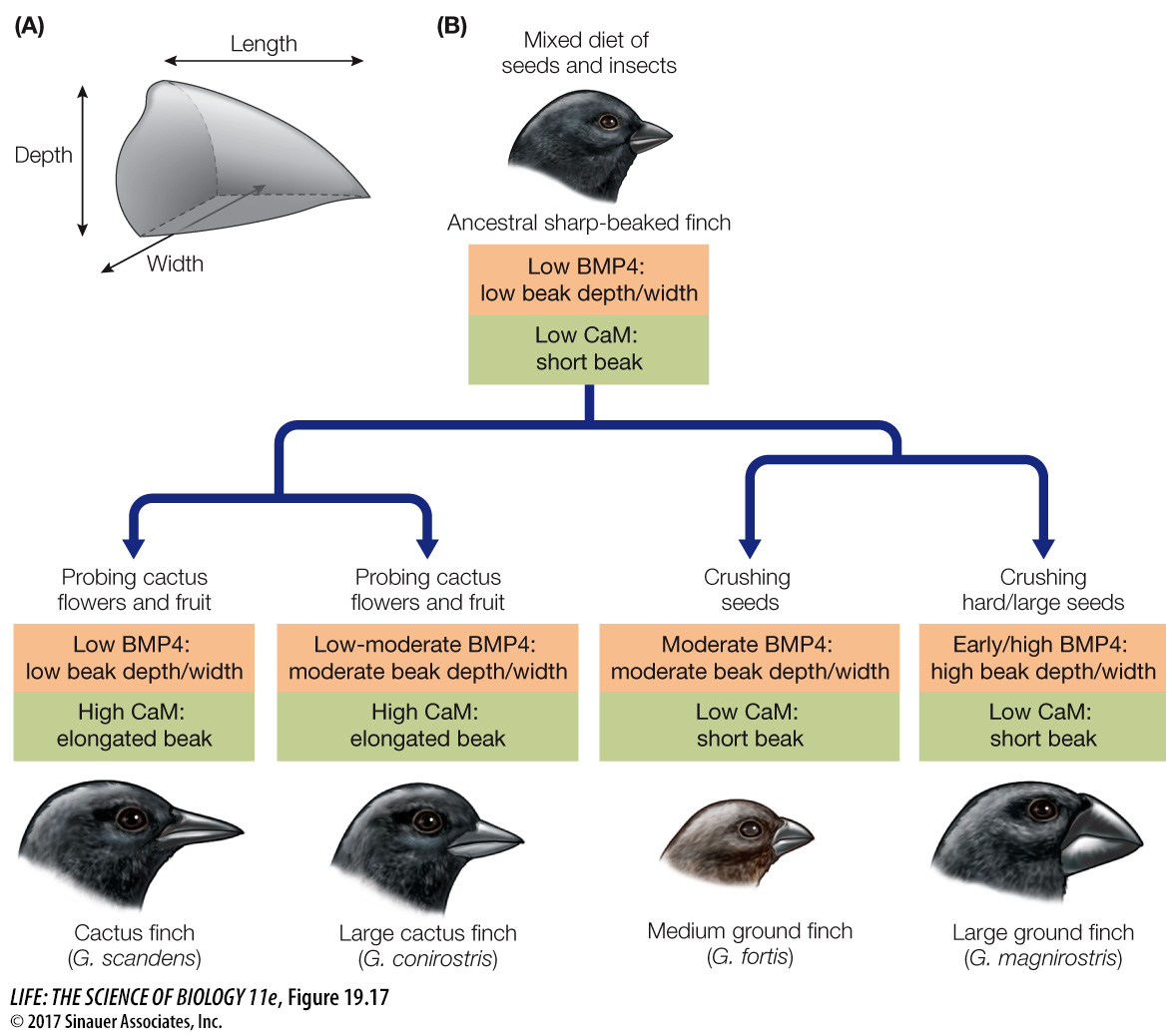
HETEROMETRY A striking example of heterometry (“different measure”) is beak development in Galápagos finches, in which the beaks of some species are large and robust for cracking seeds and in other species are thin and long for probing for food. When he visited these islands, Charles Darwin wrote that “seeing this gradation and diversity of structure in one small intimately related group of birds, one might really fancy that from an original paucity of birds in this archipelago, one species had been taken and modified for different ends.”
Darwin had no idea of the genetic basis for such modification. Now we do. Beak shapes are evident when birds hatch from their eggs, so they must be determined during development. The beak develops from tissues at the anterior of the embryo that will form the facial bones. Cell divisions in this embryonic tissue are controlled by signaling proteins, one of which is called bone morphogenetic protein 4 (BMP4); another is the protein calmodulin. If BMP4 is present early and in large amounts, the beak becomes broad and deep. If calmodulin is present early and in large amounts, the beak grows longer and thinner (Figure 19.17). Thus beak structure is affected by changes in protein production.
HETEROCHRONY The evolution of the giraffe’s neck provides an example of heterochrony (“different time”). Giraffes, like all mammals except manatees and sloths, have seven cervical (neck) vertebrae. So giraffes did not get longer necks by adding more vertebrae. However, the cervical vertebrae of giraffes are much longer than those of other mammals (Figure 19.18).
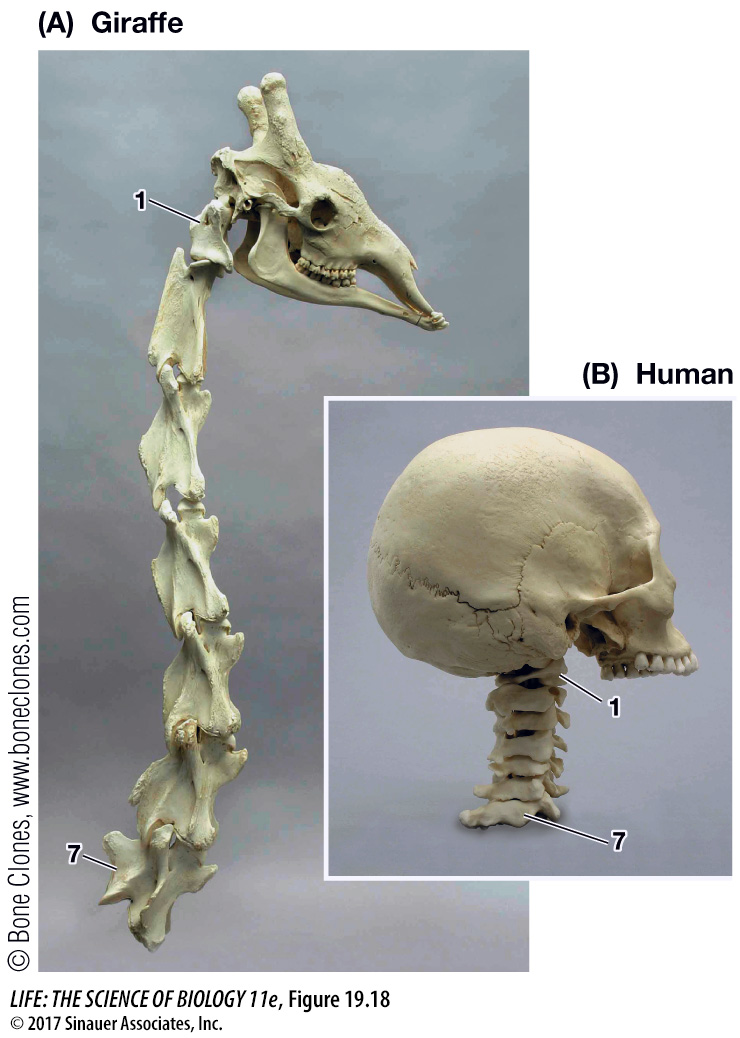
Bone growth in mammals is the result of the proliferation of cartilage-
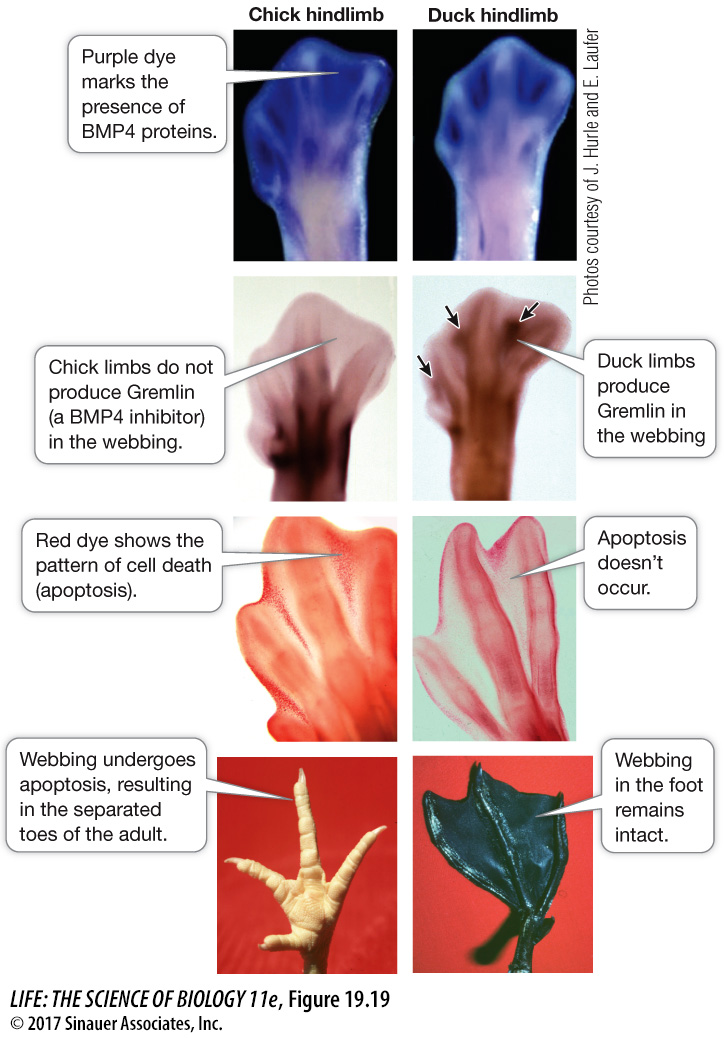
HETEROTOPY Spatial differences in the expression of a developmental gene are known as heterotopy (“different place”), exemplified by the different development of feet in ducks and chickens. The feet of all bird embryos have webs of skin that connect their toes. This webbing is retained in adult ducks (and other aquatic birds) but not in adult chickens (and other nonaquatic birds). The loss of webbing is controlled by the BMP4 signaling protein, which as we have noted is also involved in beak development (another example of use of a common genetic toolkit to produce different kinds of change).
BMP4 protein induces the cells that produce webbing to undergo apoptosis and thus eliminate the webbing between the toes. The hindlimbs of both duck and chicken embryos express the BMP4 gene in the webbing between the toes; however, they differ in expression of the Gremlin gene, which encodes a protein that inhibits BMP4 expression (Figure 19.19). In ducks, but not in chickens, Gremlin is expressed in the webbing cells and Gremlin protein inhibits BMP4 expression. With no BMP4 protein to stimulate apoptosis, a webbed foot develops. If chick hindlimbs are experimentally exposed to Gremlin during development, the adult chicken will have ducklike webbed feet.
Q: What do you predict would happen if you blocked the expression of Gremlin in the feet of duck embryos?
If Gremlin was blocked in the developing feet of duck embryos, there would not be apoptosis between the digits and so chickenlike feet would form.