Mutualistic fungi engage in relationships that benefit both partners
Certain relationships between fungi and other organisms have nutritional consequences for both partners. Two relationships of this type are symbiotic (the partners live in close, permanent contact with each other) as well as mutualistic (the relationship benefits both partners).
LICHENS A lichen is not a single organism, but rather a meshwork of at least two radically different species: a fungus and a photosynthetic alga or cyanobacterium. Recent studies suggest that an additional fungal species (a unicellular yeast) is also usually involved in this mutualism. Together the organisms that constitute a lichen can survive some of the harshest environments on Earth (although they are sensitive to poor air quality; see Key Concept 29.4). The biota of Antarctica, for example, features more than 100 times as many kinds of lichens as of plants. Relatively little experimental work has focused on lichens, perhaps because they grow so slowly—
There are nearly 30,000 described “species” of lichens, each of which is assigned the name of its multicellular fungal component. These fungal components may constitute as many as 20 percent of all fungal species. Most of them are sac fungi (Ascomycota). Some of them are able to grow independently without a photosynthetic partner, but most have never been observed in nature other than in a lichen association. The photosynthetic component of a lichen is most often a unicellular green alga, but it can be a cyanobacterium, or may even include both.
Lichens are found in all sorts of exposed habitats: on tree bark, on open soil, and on bare rock. Reindeer moss (not a moss at all, but the lichen Cladonia subtenuis) covers vast areas in Arctic, sub-
The body forms of lichens fall into three principal categories. Crustose (crustlike) lichens adhere tightly to their substrate (Figure 29.8A). Foliose (leafy) lichens are loosely attached and grow parallel to their substrate (Figure 29.8B, right). Fruticose lichens are highly branched and can grow upward like shrubs, or hang in long strands from tree branches or rocks (Figure 29.8B, left).

A cross section of a typical foliose lichen reveals a tight upper region of fungal hyphae, a layer of photosynthetic cyanobacteria or algae, a looser hyphal layer, and finally hyphal rhizoids that attach the entire structure to its substrate (Figure 29.9). The meshwork of fungal hyphae takes up mineral nutrients needed by the photosynthetic cells and also holds water tenaciously, providing a suitably moist environment. The fungus obtains fixed carbon from the photosynthetic products of the algal or cyanobacterial cells.
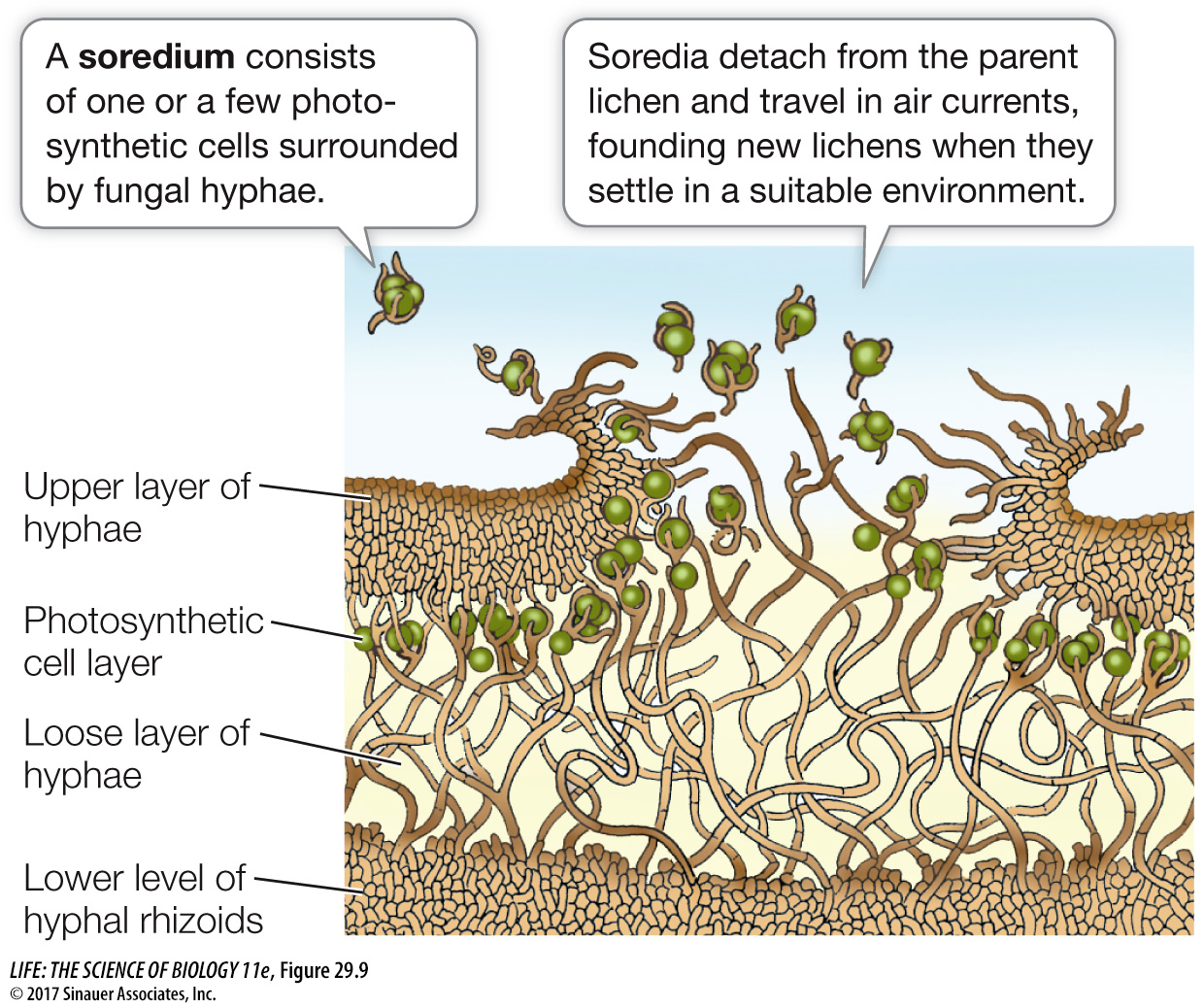
Within the lichen, fungal hyphae are tightly pressed against the photosynthetic cells and sometimes invade them without breaching the cell membrane (similar to the haustoria in parasitic fungi; see Figure 29.5). The photosynthetic cells not only survive these intrusions but continue to grow. Algal cells in a lichen “leak” photosynthetic products at a greater rate than do similar cells growing on their own, and photosynthetic cells taken from lichens grow more rapidly on their own than when associated with a fungus. On the basis of these observations, we could consider lichen fungi to be parasitic on their photosynthetic partners. In many places where lichens grow, however, the photosynthetic cells could not grow at all on their own.
Lichens can reproduce simply by fragmentation of the vegetative body (the thallus) or by means of specialized structures called soredia (singular soredium). Soredia consist of one or a few photosynthetic cells bound by fungal hyphae. They become detached from the lichen, are dispersed by air currents, and upon arriving at a favorable location, develop into a new lichen thallus (see Figure 29.9). Alternatively, the fungal partner may go through its sexual reproductive cycle, producing haploid spores. When these spores are discharged, however, they disperse alone, unaccompanied by the photosynthetic partner.
Lichens are often the first colonists on new areas of bare rock. They get most of the mineral nutrients they need from the air and rainwater, augmented by minerals absorbed from dust. A lichen begins to grow shortly after a rain, as it begins to dry. As it grows, the lichen acidifies its environment slightly, and this acidity contributes to the slow breakdown of rocks, an early step in soil formation. With further drying, the lichen’s photosynthesis ceases. The water content of the lichen may drop to less than 10 percent of its dry weight, at which point it becomes highly insensitive to extremes of temperature.
MYCORRHIZAE Many vascular plants depend on a symbiotic association with fungi. This ancient association between plants and fungi was critical to the successful exploitation of the terrestrial environment by plants. Unassisted, the root hairs of many plants often do not take up enough water or minerals to sustain growth. However, the roots of such plants usually do become infected with fungi, forming an association called a mycorrhiza (plural mycorrhizae). There are two types of mycorrhizae, distinguished by whether or not the fungal hyphae penetrate the plant cell walls.
In ectomycorrhizae, the fungus wraps around the root tip, and its mass is often as great as that of the root itself (Figure 29.10A). The fungal hyphae penetrate the root and wrap around individual root cells but do not penetrate the cell walls. An extensive web of hyphae penetrates the soil in the area around the root, so that up to 25 percent of the volume near the root may be fungal hyphae. The hyphae attached to the root increase the surface area for the absorption of water and minerals, and the mass of hyphae in the soil acts like a sponge to hold water in the neighborhood of the root. Infected roots are short, swollen, and club-
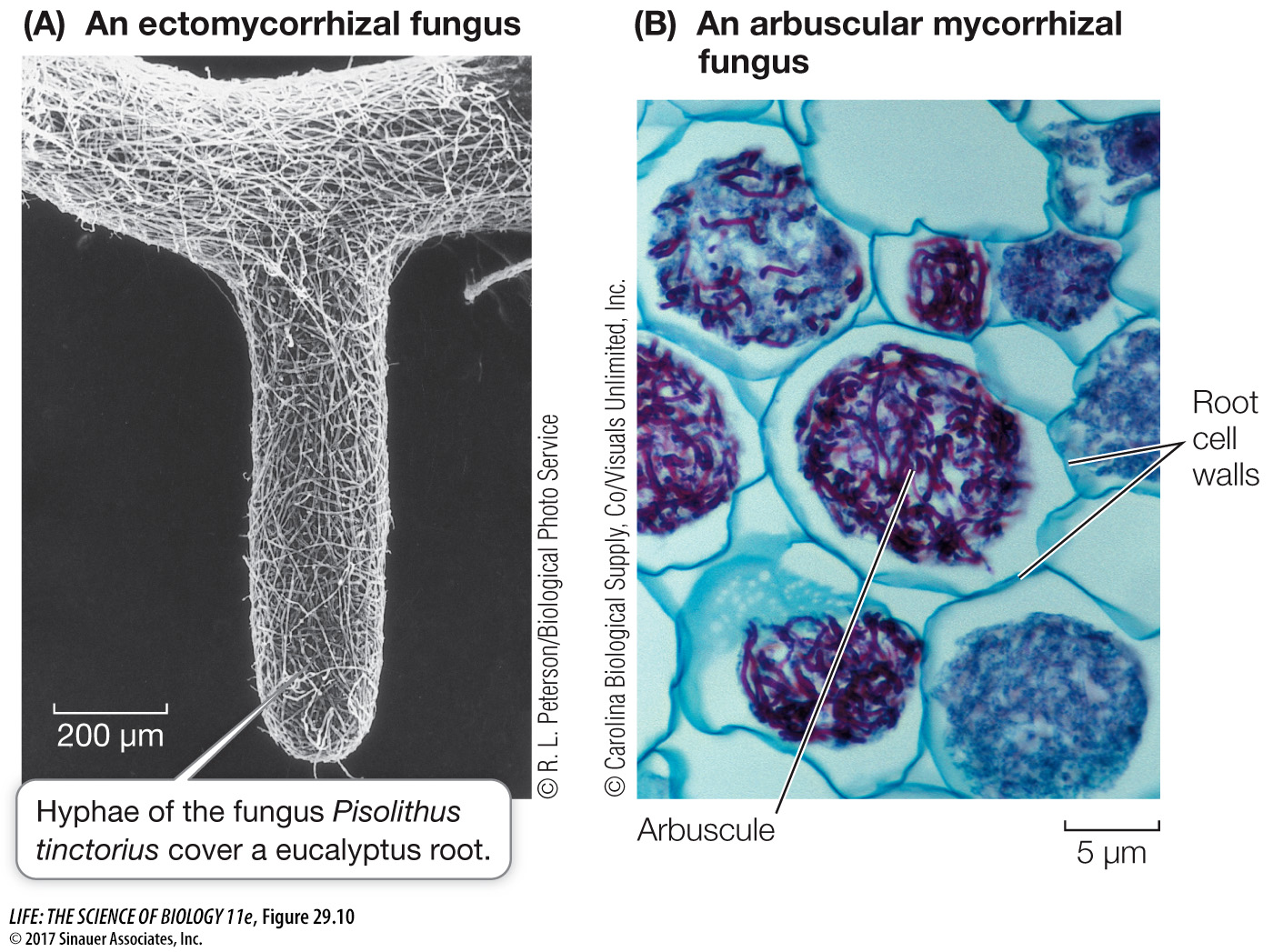
The fungal hyphae of *arbuscular mycorrhizae enter the root and penetrate the cell walls of the root cells, forming arbuscular (treelike) structures inside the cell wall but outside the cell membrane. These structures, like the haustoria of parasitic fungi and the contact regions of fungal hyphae and photosynthetic cells in lichens, become the primary site of exchange between plant and fungus (Figure 29.10B). As in the ectomycorrhizae, the fungus forms a vast web of hyphae leading from the root surface into the surrounding soil.
*connect the concepts Mycorrhizae can vastly increase the root surface area in contact with essential nutrients in the soil. The importance of arbuscular mycorrhizae in plant nutrition is described in detail in Key Concept 35.4.
The mycorrhizal association is important to both partners. The fungus obtains needed organic compounds, such as sugars and amino acids, from the plant. In return, the fungus, because of its very high surface area-
Plants that have active arbuscular mycorrhizae typically are a deeper green and may resist drought and temperature extremes better than plants of the same species that have little mycorrhizal development. Attempts to introduce some plant species to new areas have failed until some of soil from the native area (presumably containing the fungus necessary to establish mycorrhizae) was provided. Trees without ectomycorrhizae do not grow well in the absence of abundant nutrients and water, so the health of our forests depends on the presence of ectomycorrhizal fungi. Many agricultural crops require inoculation of seeds with appropriate mycorrhizal fungi prior to planting. Without these fungi, the plants are unlikely to grow well, or in some cases at all. Certain plants that live in nitrogen-