Many stems and roots undergo secondary growth
As you have seen, the roots and stems of some eudicots develop a secondary plant body, the tissues of which include wood and bark. These tissues are derived by secondary growth from the two lateral meristems:
The vascular cambium is a cylindrical layer of tissue consisting predominantly of elongated cells that divide frequently. It supplies the cells of the secondary xylem and secondary phloem, which eventually become wood and bark, respectively.
The cork cambium produces mainly waxy-
walled protective cells. It supplies some of the cells that become bark.
investigating life
Understanding the Synthesis and Transport of Cyanogenic Glycosides
experiment
Original Paper: Jørgensen, K. et al. 2005. Cassava plants with a depleted cyanogenic glycoside content in leaves and tubers. Distribution of cyanogenic glycosides, their site of synthesis and transport and blockage of biosynthesis by RNA interference technology. Plant Physiology 139: 363–
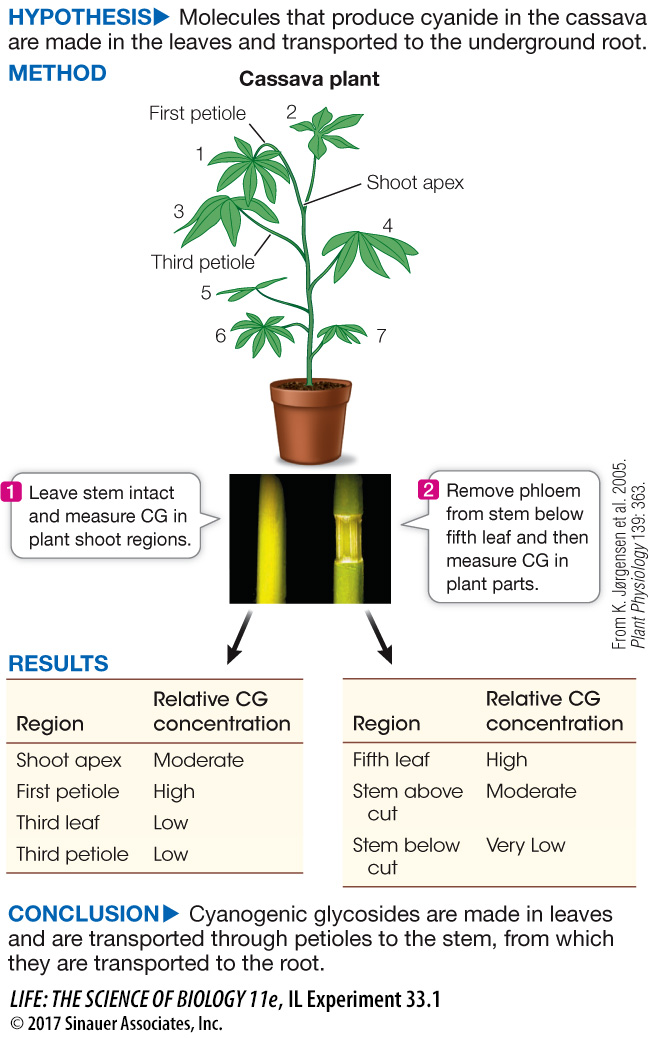
As described in the chapter opening, cassava is the most important root crop in the world. Its use as food is challenging, because the plant produces molecules that can be transformed into toxic cyanide when the tissues are disrupted. These cyanogenic glycosides (CGs) are important in defending the plant against animals that eat it, including humans. For the millions of people who eat cassava, careful processing is needed to avoid poisoning. Unfortunately, the processing methods lower the overall nutritional value of the food. Understanding how CGs are made and end up in the fleshy root is an important step in possibly using genetics to produce a crop that does not make CGs. At the Royal Veterinary and Agricultural University in Copenhagen, a team led by Birger Møller investigated the synthesis and transport of CGs in cassava plants.
work with the data
QUESTIONS
1.
Cyanogenic glycosides in plant organs were measured in an intact stem of a cassava plant and in one in which the phloem had been cut below the fifth leaf. The results are shown in the graphs. What can you conclude about the source of CG?
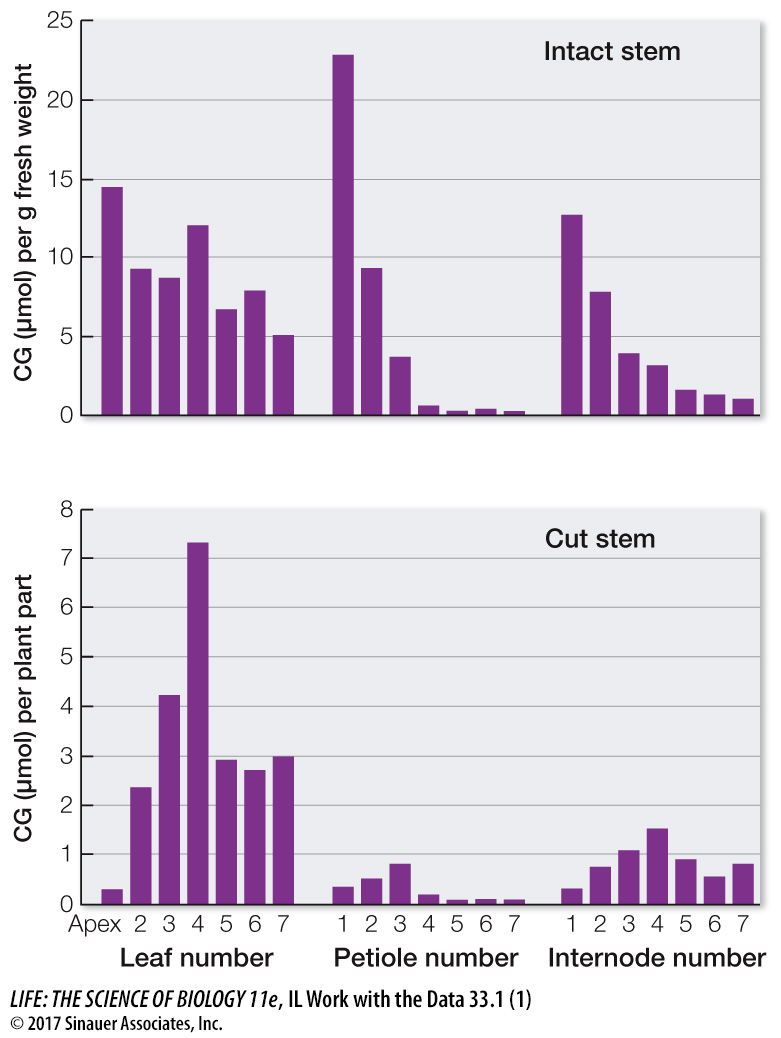
CG appears to be highest in the upper leaves and shoot, where it is made. The lower leaves and stems contain less CG, indicating that CG is transported there on the way to other plant organs.
2.
When a stem is girdled, it is cut carefully so that the epidermal, cortical, and outer vascular tissues (phloem) are removed. Using intact plants, the biologists girdled the stem of a cassava plant. After 2 days, they measured CG in various regions on the stem. The results for CG levels after girdling between the fifth and sixth nodes are shown in the table. What can you conclude about the source and transport of CG? Why was the level in the sixth leaf (below the girdle) so high?
| Region | CG level (μmol) |
|---|---|
| Fifth leaf | 59 |
| Stem above cut | 15 |
| Stem below cut | 0.2 |
| Sixth leaf | 55 |
The fifth leaf accumulated CG, which was not transported down the plant vascular system because of girdling. There was some residual accumulation in the sixth leaf that had probably arrived before the girdling. The stem below the sixth leaf had very low CG, indicating the transport was blocked.
A similar work with the data exercise may be assigned in LaunchPad.
In cold parts of the world, each year, deciduous trees lose their leaves and have bare branches and twigs over the winter (Figure 33.17). The apical meristems of the twigs are enclosed in buds protected by bud scales. When the buds begin to grow in spring, the scales fall away, leaving scars that show where the bud was. These scars allow us to identify the parts of the twig from each year’s growth. The dormant twig shown in Figure 33.17 is the product of both primary and secondary growth. The buds consist entirely of primary tissues.
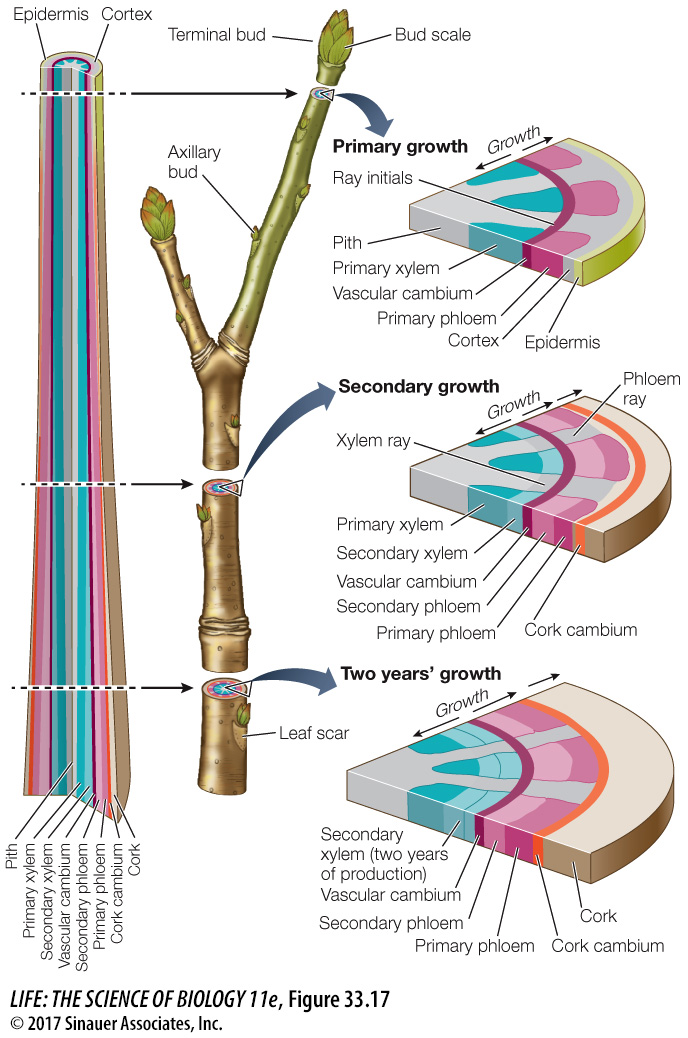
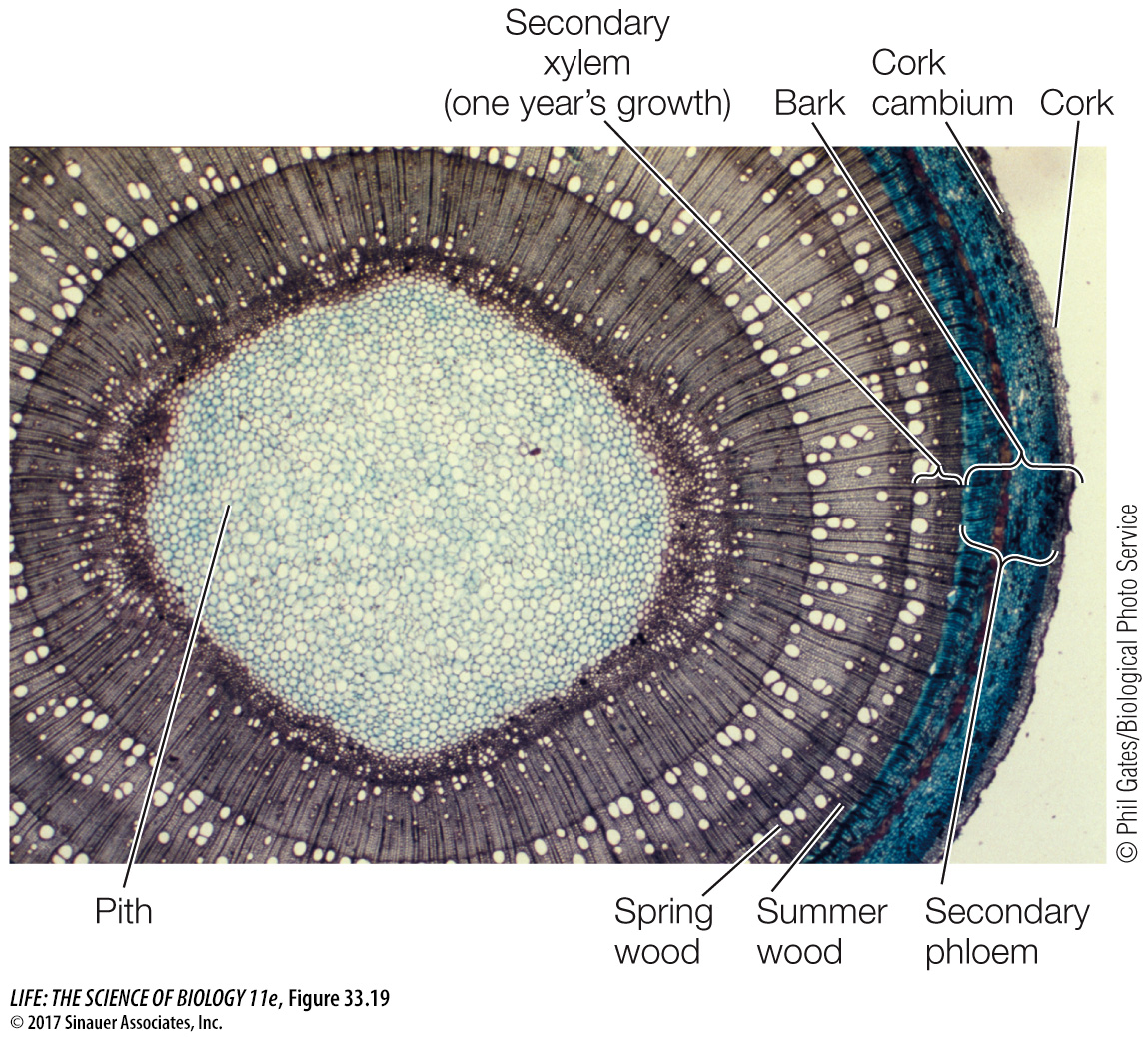
The vascular cambium is initially a single layer of cells lying between the primary xylem and the primary phloem within the vascular bundles. The root or stem increases in diameter when the cells of the vascular cambium divide, producing secondary xylem cells toward the inside of the root or stem and producing secondary phloem cells toward the outside. In the stem, cells in the pith rays between the vascular bundles also divide, forming a continuous cylinder of vascular cambium running the length of the stem. This cylinder, in turn, gives rise to complete cylinders of secondary xylem (the wood) and secondary phloem, which contributes to the bark. It also produces vascular rays for lateral transport, a structure not found in primary xylem and phloem. Therefore the vascular cambium produces vessel elements, tracheids, parenchyma cells, and supportive fibers in the secondary xylem; and sieve tube elements, companion cells, fibers, and parenchyma cells in the secondary phloem.
Animation 33.1 Secondary Growth: The Vascular Cambium
As secondary growth of stems or roots continues, the expanding vascular tissue stretches and breaks the epidermis and the outer layers of the cortex, which ultimately flake away. Before these dermal tissues are broken away, cells lying near the surface of the secondary phloem begin to divide, forming a cork cambium. This meristematic tissue produces layers of cork, a protective tissue composed of cells with thick walls waterproofed with suberin. The cork soon becomes the outermost tissue of the stem or root (see Figure 33.17). Without the activity of the cork cambium, the sloughing off of the outer primary tissues would expose the plant to potential damage, such as excessive water loss or invasion by microorganisms. Sometimes the cork cambium produces cells toward the inside as well as the outside; these cells constitute a tissue known as the phelloderm.
The cork cambium, cork, and phelloderm constitute the secondary dermal tissue called periderm. As the vascular cambium continues to produce secondary vascular tissue, these corky layers are lost, but the continuous formation of new cork cambia in the underlying secondary phloem gives rise to new corky layers. The periderm and the secondary phloem—
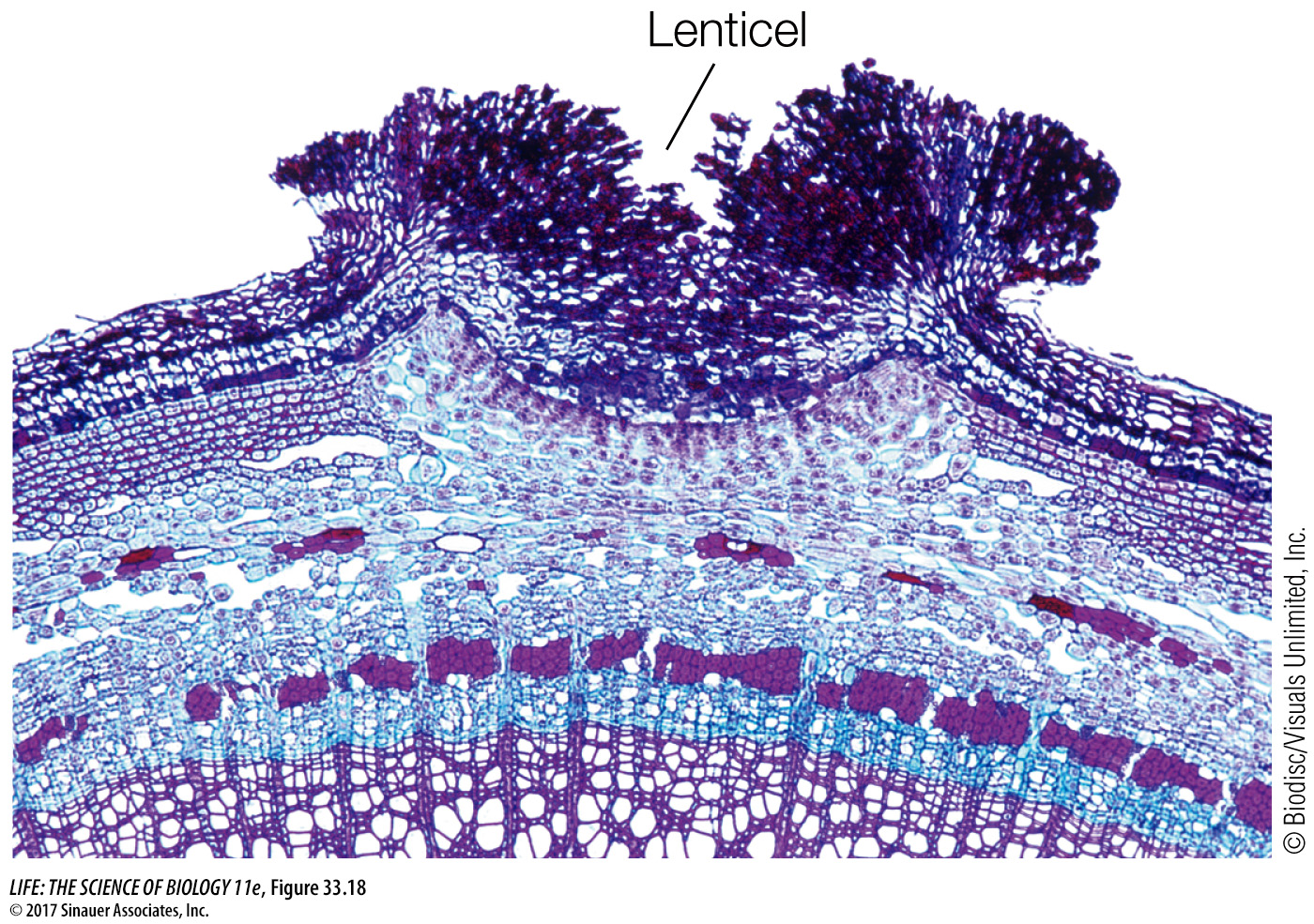
When periderm forms on stems or roots, the underlying tissues still need to release carbon dioxide and take up oxygen for cellular respiration. Spongy regions in the periderm called lenticels can allow such gas exchange (Figure 33.18).
You may be curious about cork, which is used as stoppers in bottles of wine and in flasks in the chemistry lab. Cork is harvested from a tree, the cork oak (Quercus suber, see photo below), which is grown for that purpose mainly in Portugal. The tree lives more than 100 years, and cork is first harvested when the tree is 25 years old. The outer bark is removed first, carefully exposing the cork layers, which are then removed without damaging the living tissues. The cork cambium then produces more cork, so a tree can be harvested every 10 years.

Cross sections of most tree trunks (stems) in temperate-
Only eudicots and other non-