The pressure flow model accounts for translocation in the phloem
The observations we just described have led to the pressure flow model as an explanation for translocation in the phloem (Figure 34.10). According to this model, sucrose is actively transported at a source into companion cells, from which it flows through plasmodesmata into the sieve tube elements. This gives those cells a higher sucrose concentration than the surrounding cells (a more negative solute potential), and water therefore enters the sieve tube elements from the xylem by osmosis. The entry of this water increases turgor pressure (causes a more positive pressure potential) at the source end of the sieve tube, so that the entire fluid content of the sieve tube is pushed toward the sink end of the tube—
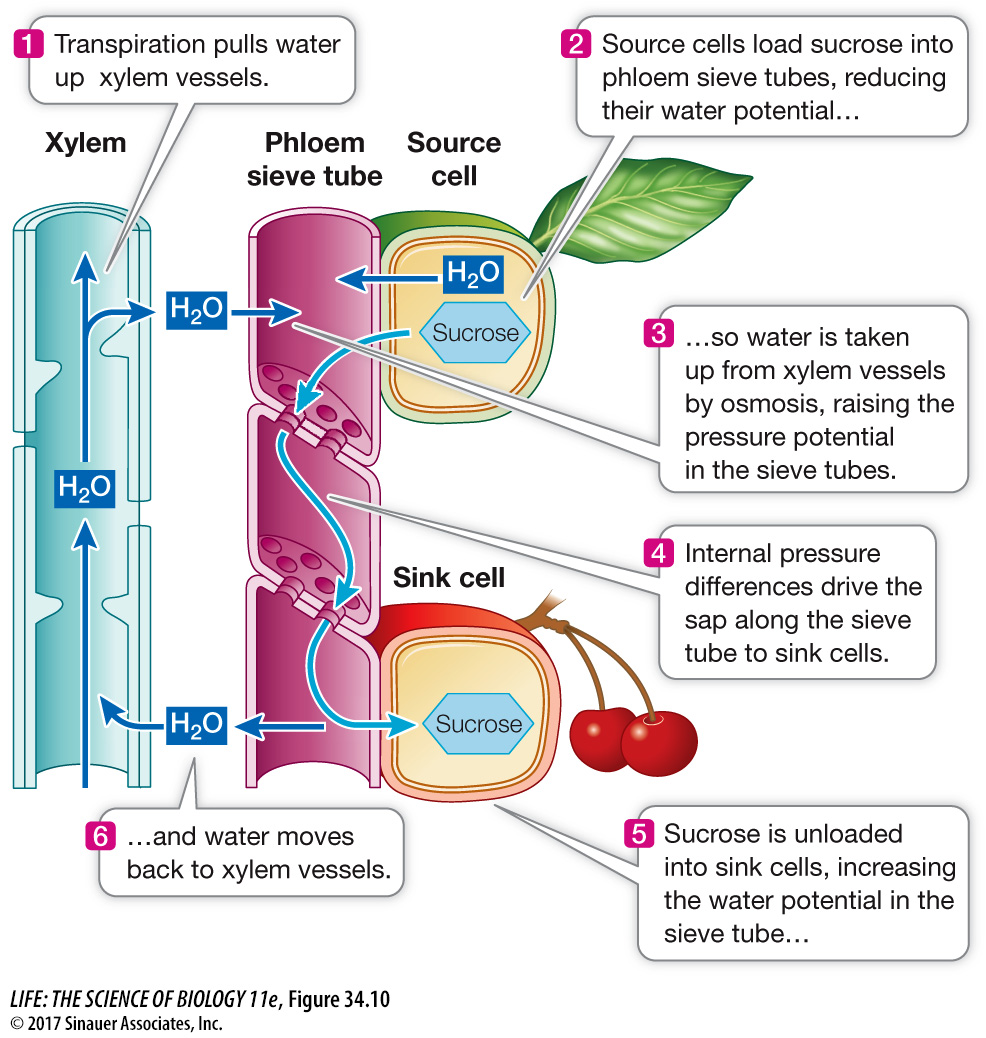
Animation 34.2 The Pressure Flow Model
Two steps in phloem translocation require metabolic energy:
Loading: Transport of sucrose and other solutes from sources into companion cells and then into the sieve tubes.
Unloading: Transport of solutes from the sieve tubes into sinks. The need for *metabolic energy is the reason why phloem transport, unlike xylem transport, requires living cells.
*connect the concepts Interactions among biochemical pathways (such as photosynthesis, cellular respiration, and the Calvin cycle) that produce carbohydrates that fuel processes such as translocation are discussed in Key Concept 10.5.
Sucrose and other solutes can move from the mesophyll cells into the phloem by either an apolastic pathway or a symplastic pathway. In the apolastic pathway, sucrose and other solutes leave the mesophyll cells and enter the apoplast before they reach the sieve tube elements. Specific sugars and amino acids are then actively transported into cells of the phloem. Because the solutes cross at least one selectively permeable membrane in the apoplastic pathway, selective transport can be used to regulate which specific substances enter the phloem. In the symplastic pathway, the solutes remain within the symplast all the way from the mesophyll cells to the sieve tube elements. Because no membranes are crossed in the symplastic pathway, a mechanism that does not involve membrane transport is used to load sucrose into the phloem.
In sink regions, the solutes are actively transported out of the sieve tube elements and into the surrounding tissues. This unloading serves several purposes: it helps maintain the gradient of solute potential, and hence of pressure potential, in the sieve tubes; it helps build up high concentrations of carbohydrates in storage organs, such as developing fruits and seeds; and it moves nutrients to parts of the plant that are growing rapidly, such as shoots that form after the winter.
Table 34.2 contrasts the pressure flow model of translocation in the phloem with the transpiration–
| Xylem | Phloem | |
|---|---|---|
| Driving force for bulk flow | Transpiration from leaves | Active transport of sucrose at source and sink |
| Site of bulk flow | Nonliving vessel elements and tracheids | Living sieve tube elements |
| Pressure potential in sap | Negative (pull from top; tension) | Positive (push from source; pressure) |
The control of phloem sap travel from source to sink has great importance for people. The parts of plants that we use for food, such as seeds (e.g., the rice in the opening story), fruits, and storage organs, are mostly sinks. Increasing the flow of sucrose into these organs as they develop can increase food production in crop plants.