Sleep and dreaming are reflected in electrical patterns in the cerebral cortex
A dominant feature of behavior is the daily cycle of sleep and waking. All birds and mammals, probably all other vertebrates, and also many invertebrates, sleep. We humans spend one-third of our lives sleeping, yet we do not know why or how. We do know, however, that we need to sleep. Loss of sleep impairs alertness and performance. Many people in our society—certainly most college students—are chronically sleep-deprived. Accidents and serious mistakes that endanger lives can be attributed to impaired alertness caused by lack of sleep. Insomnia (difficulty in falling or staying asleep) is one of the most common medical complaints.
THE ELECTROENCEPHALOGRAM A common tool of sleep researchers is the electroencephalogram, or EEG. Rather than recording the activity of single neurons, the EEG characterizes activity in huge numbers of neurons. EEG electrodes are much larger than the fine electrodes used to detect single cell activity. Placed at different locations on the head and scalp (Figure 46.11A), EEG electrodes record fluctuations in the electric potential differences between electrodes over time. These differences reflect the electrical activity of the neurons in the brain regions under the electrodes, primarily regions of the cerebral cortex. Usually the electrical activity of one or more skeletal muscles is also recorded; this record is called an electromyogram (EMG). Movements of the eyes are recorded as an electroocculogram (EOG).
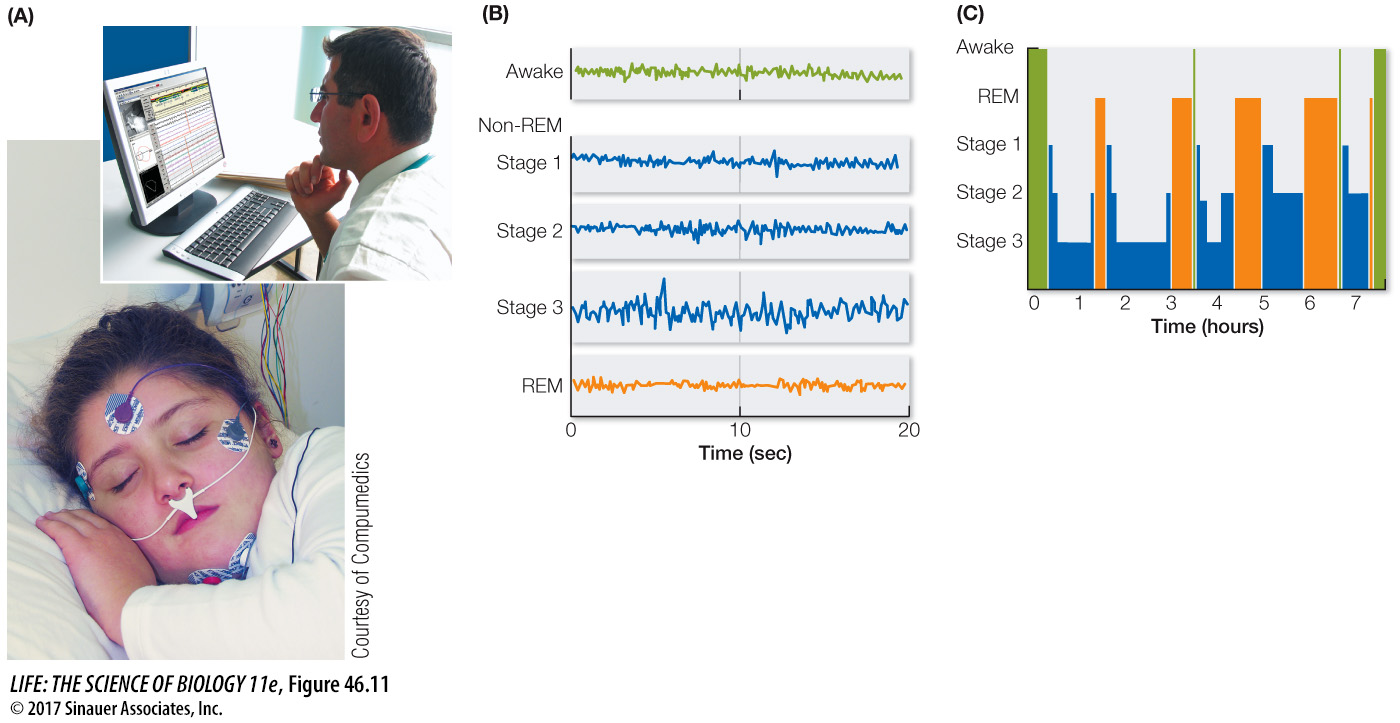
Figure 46.11 Stages of Sleep (A) Electrical activity in the cerebral cortex is detected by electrodes that are placed on the head and record changes in voltage between the electrodes through time. (B) The resulting record is an electroencephalogram, or EEG. (C) Humans cycle through different stages of sleep throughout the night.
EEG, EMG, and EOG patterns reveal the transition from being awake to being asleep. They also reveal that there are different states of sleep. In mammals other than humans, two major sleep states are easily distinguished: slow-wave sleep and rapid eye movement (REM) sleep. Slow-wave sleep gets its name from the high-amplitude, slow-frequency waves in the EEG. REM sleep gets its name from jerky movements of the eyeballs that occur during this state. In humans, sleep states are characterized as non-REM sleep and REM sleep. Human non-REM sleep is divided into three stages.
When you fall asleep, you first enter stage 1 non-REM sleep, which then progresses through stage 2 to stage 3 (Figure 46.11B). Stage 3 is deep, restorative, slow-wave sleep. This first full cycle of non-REM sleep is followed by an episode of REM sleep. Throughout the night you experience four or five cycles of non-REM and REM sleep (Figure 46.11C). About 80 percent of your sleep is non-REM sleep. The most vivid dreams and nightmares occur during the 20 percent of sleep that is REM sleep.
Page 995
CELLULAR CHANGES DURING SLEEP When we are awake, several nuclei (groups of neurons) in the brainstem reticular-activating system are continuously active. Axons from neurons in these nuclei extend to the thalamus and throughout the cerebral cortex, where they release depolarizing neurotransmitters (acetylcholine, norepinephrine, and serotonin). These broadly distributed neurotransmitters keep the resting potential of the neurons of the thalamus and cortex close to threshold and sensitive to synaptic inputs, thereby maintaining the responsiveness of the brain that characterizes being awake.
With the onset of sleep, activity in these brainstem nuclei decreases, and their axon terminals release less neurotransmitter. With the withdrawal of the depolarizing neurotransmitters, the resting potentials of the cells of the thalamus and cortex become more negative (hyperpolarized), and the cells are less sensitive to excitatory synaptic input. Their processing of information is inhibited, and consciousness is lost. You have probably experienced this as “nodding off.” As you enter non-REM sleep, cells of the cortex begin to fire action potentials in bursts. The synchronization of these bursts over broad areas of cerebral cortex results in the EEG slow-wave pattern that characterizes deep non-REM sleep.
At the transition from non-REM to REM sleep, dramatic changes occur. Some of the brainstem nuclei that were inactive during non-REM sleep become active again, causing a general depolarization of cortical neurons. Thus in REM sleep the synchronized bursts of firing cease, and the EEG resembles that of the awake brain. Because the resting potentials of the neurons return to near threshold levels, the cortex can process information, and vivid dreams occur.
So why don’t we act out our dreams? During REM sleep the brain inhibits both afferent (sensory) and efferent (motor) pathways; we are paralyzed during REM sleep. Limb twitches and the jerky eye movements are motor signals breaking through the inhibition. The bizarre nature of dreams may be due to the lack of sensory feedback to the cortex from the body and the outside world. In other words, a functioning cortex is out of touch with reality. The function of muscle paralysis during REM sleep may be to prevent the acting out of dreams.
Knowing the cellular mechanisms of sleep has not yet led to an understanding of its function. Many questions remain. Why do we have two sleep states with very different neurophysiological characteristics? Why does non-REM sleep always occur first? Why do the two states cycle during the rest phase? We know sleep is essential for life, but we don’t know why. One set of hypotheses is that sleep is necessary for the maintenance and repair of neural connections and for the neural changes involved in learning and memory—and possibly forgetting. These hypotheses are supported by many experiments showing that performance of a learned task or recall of declarative information on the day following training is impaired if sleep is prevented, and is best following a good night’s sleep.