Materials are exchanged in capillary beds by filtration, osmosis, and diffusion
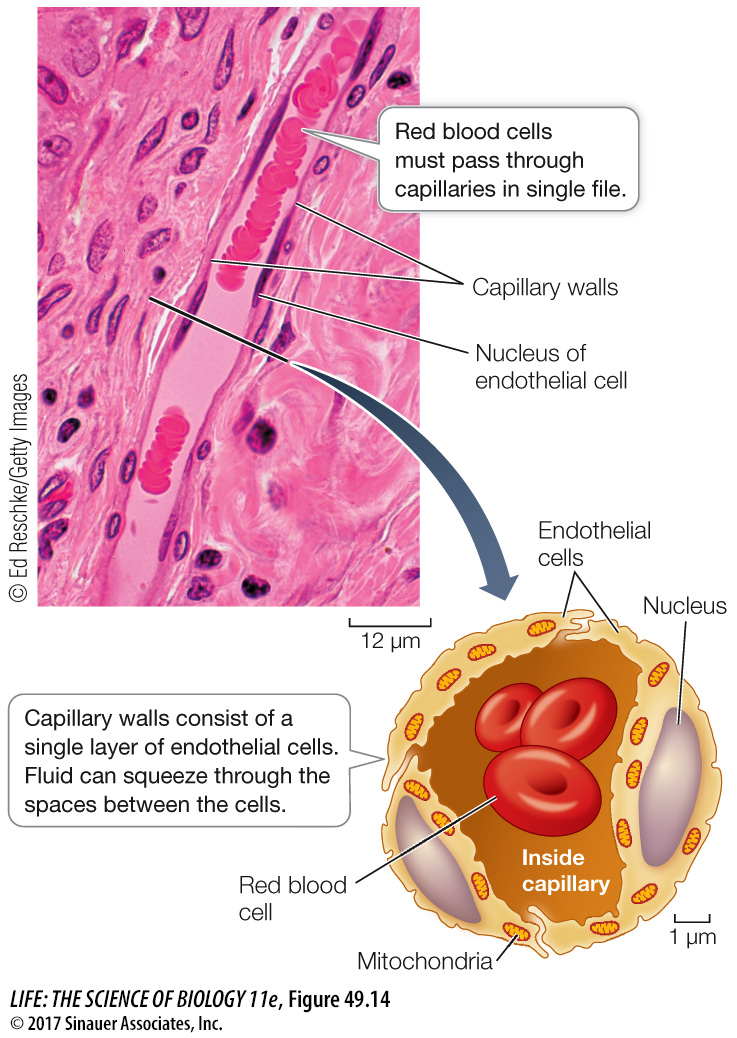
Beds of capillaries lie between arterioles and venules (Figure 49.13B). Most cells are not more than a few cell diameters away from a capillary. The needs of cells are served by the exchange of materials between blood and interstitial fluid across the capillary walls. Capillary walls are thin and permeable to water and many solutes. Also, blood flows slowly through capillaries, allowing time for exchange.
It may seem strange that blood flows through the large arteries rapidly at high pressures, but when it reaches the small capillaries the pressure and rate of flow decrease (Figure 49.13C). When you constrict the opening of a garden hose, the pressure in the hose and the velocity of the water spraying out increase. But keep in mind that the arteries branch into many arterioles that serve huge numbers of capillaries. Even though each capillary has a diameter so small that red blood cells pass through in single file, there are so many capillaries that their total cross-
Capillary walls consist of a single layer of endothelial cells (Figure 49.14). Capillaries are permeable to water, some ions, and some small molecules but not to large molecules such as proteins, and certainly not to red blood cells. At the arterial (high pressure) end, blood pressure squeezes water and small solutes out through spaces between the cells of the capillary walls into the surrounding intercellular space.
Media Clip 49.1 Capillary Flow: A Tight Squeeze
Why don’t water and small-
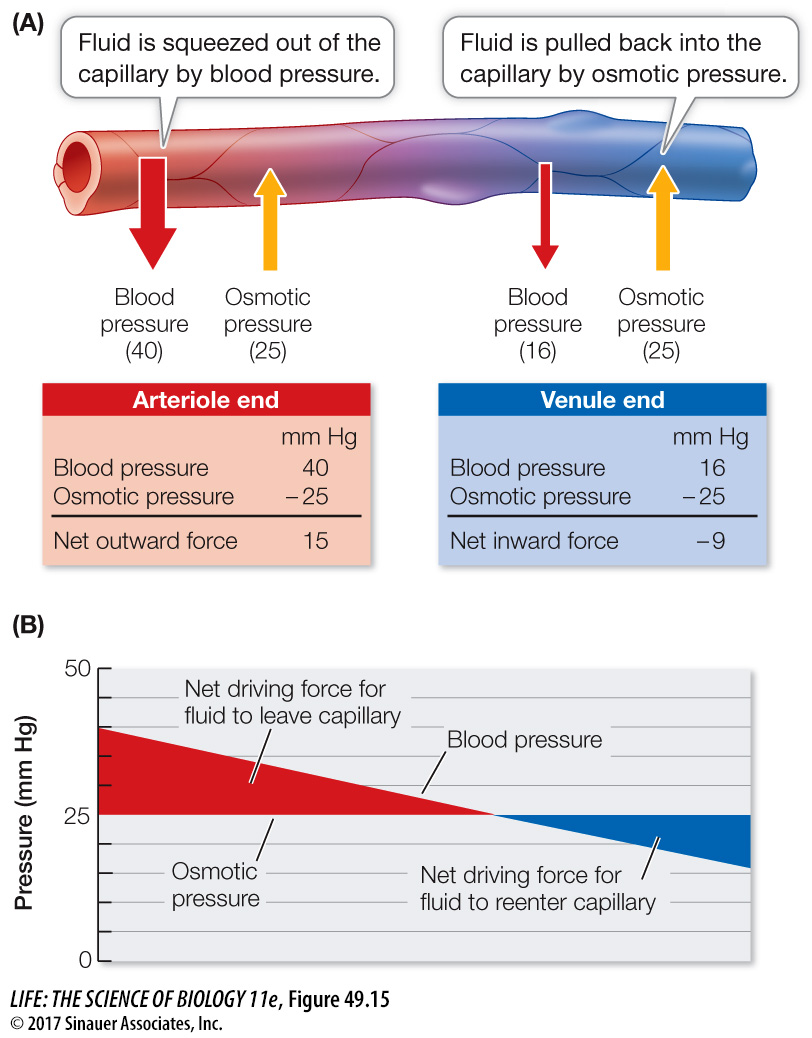
Blood hydrostatic pressure squeezes water and small solutes out of the capillaries.
Blood osmotic pressure pulls water back into the capillaries.
Blood hydrostatic pressure is high at the arterial end of a capillary bed and steadily drops as blood moves toward the venous end (Figure 49.15). Blood osmotic pressure is due to the large protein molecules that cannot leave the capillaries, and it is relatively constant along the capillaries. As long as the blood hydrostatic pressure is above the blood osmotic pressure, fluid leaves the capillaries. At the venule end of most capillaries, blood hydrostatic pressure falls below blood osmotic pressure, so fluid returns to the capillaries. The actual numbers for a normal capillary bed in a resting person suggest that there would be a slight net loss of fluid to the intercellular spaces. This loss, about 4 liters per day, percolates between cells as the interstitial fluid before it returns to the venous blood via the lymphatic system, which we will discuss later in this chapter.
Recent research suggests that in addition to large protein molecules in the blood, bicarbonate ions (HCO3–) may also contribute significantly to pulling water back into the capillaries. The CO2 produced by cellular metabolism diffuses into the endothelial cells lining the capillaries, where it is converted into HCO3– and released into the plasma. When an individual is at rest, the increasing HCO3– concentration can cause the blood osmotic pressure at the venous end to be 30 mm Hg higher than at the arterial end, and during strenuous exercise this difference can be much higher. Thus it appears that CO2 and HCO3– are major factors that pull water back into the capillaries.
All capillaries are permeable to O2, CO2, and small ions. Lipid-
The capillaries of the brain are a special case, being rather impermeable and wrapped by glia. Not much can pass through them other than lipid-