Self-Splicing Group II Introns Provide Clues to the Evolution of snRNAs
Under certain unphysiological in vitro conditions, pure preparations of some RNA transcripts slowly splice out introns in the absence of any protein. This observation led to the recognition that some introns are self-splicing. Two types of self-splicing introns have been discovered: group I introns, present in nuclear rRNA genes of protozoans, and group II introns, present in protein-coding genes and some rRNA and tRNA genes in mitochondria and chloroplasts of plants and fungi. Discovery of the catalytic activity of self-splicing introns revolutionized our thinking about the functions of RNA. As discussed in Chapter 5, RNA is now known to catalyze peptide-bond formation during protein synthesis in ribosomes. Here we discuss the probable role of group II introns, now found only in mitochondrial and chloroplast DNA, in the evolution of snRNAs; the functioning of group I introns is considered in the later section on rRNA processing.
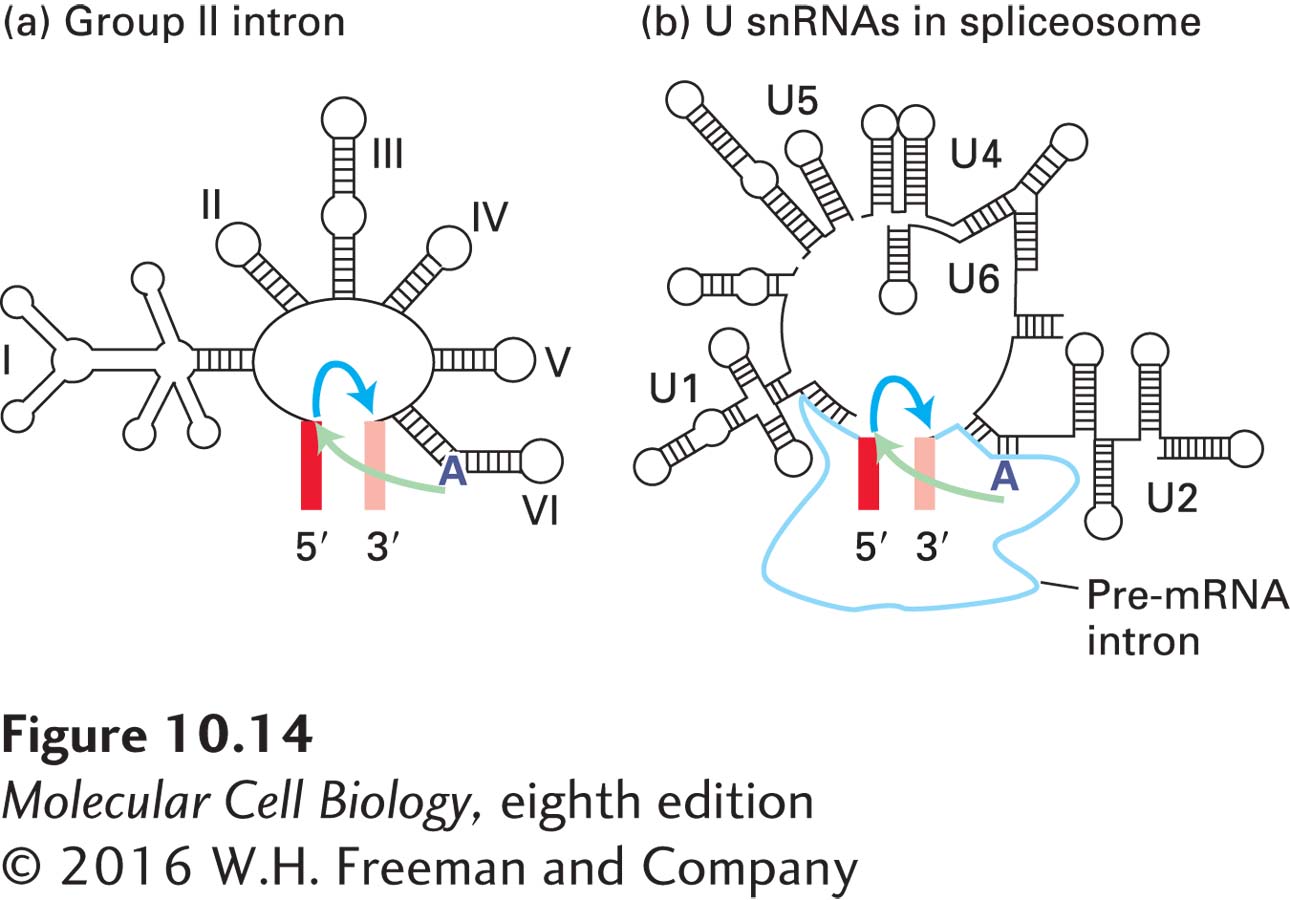
Even though their precise sequences are not highly conserved, all group II introns fold into a conserved, complex secondary structure containing numerous stem-loops (Figure 10-14a). Self-splicing by a group II intron occurs via two transesterification reactions involving intermediates and products analogous to those found in nuclear pre-mRNA splicing. The mechanistic similarities between group II intron self-splicing and spliceosomal splicing led to the hypothesis that snRNAs function analogously to the stem-loops in the secondary structure of group II introns. According to this hypothesis, snRNAs interact with 5′ and 3′ splice sites of pre-mRNAs and with one another to produce a three-dimensional RNA structure that is functionally analogous to that of group II self-splicing introns (Figure 10-14b).
FIGURE 10-14Comparison of group II self-splicing introns and the spliceosome. These schematic diagrams compare the secondary structures of (a) group II self-splicing introns and (b) U snRNAs present in the spliceosome. The first transesterification reaction is indicated by light green arrows; the second reaction, by blue arrows. The branch-point A is boldfaced. The similarity in these structures suggests that the spliceosomal snRNAs evolved from group II introns, and that the trans-acting snRNAs are functionally analogous to the corresponding domains in group II introns. The colored bars flanking the introns in (a) and (b) represent exons. See P. A. Sharp, 1991, Science254:663.
Page 430
An extension of this hypothesis is that introns in ancient pre-mRNAs evolved from group II self-splicing introns through the progressive loss of internal RNA structures, which concurrently evolved into trans-acting snRNAs that perform the same functions. Support for this type of evolutionary model comes from experiments with group II intron mutants in which domain V and part of domain I are deleted. RNA transcripts containing such mutant introns are defective in self-splicing, but when RNA molecules equivalent to the deleted regions are added to the in vitro reaction, self-splicing occurs. This finding demonstrates that these domains in group II introns can be trans-acting, like snRNAs.
The similarity in the mechanisms of group II intron self-splicing and of spliceosomal splicing of pre-mRNAs also suggests that the splicing reaction is catalyzed by the snRNA, not the protein, components of spliceosomes. Although group II introns can self-splice in vitro at elevated temperatures and Mg2+ concentrations, under in vivo conditions, proteins called maturases, which bind to group II intron RNA, are required for rapid splicing. Maturases are thought to stabilize the precise three-dimensional interactions of the intron RNA required to catalyze the two splicing transesterification reactions. By analogy, snRNP proteins in spliceosomes are thought to stabilize the precise geometry of snRNAs and intron nucleotides required to catalyze pre-mRNA splicing.
The evolution of snRNAs may have been an important step in the rapid evolution of higher eukaryotes. As sequences involved in self-splicing were lost from introns and their functions supplanted by trans-acting snRNAs, the remaining intron sequences would have become free to diverge. This in turn probably facilitated the evolution of new genes through exon shuffling, since there would be few constraints on the sequences of new introns generated in the process (see Figures 8-18 and 8-19). It also permitted the increase in protein diversity that results from alternative RNA splicing and an additional level of gene control resulting from regulated RNA splicing.