A Cascade of Regulated RNA Splicing Controls Drosophila Sexual Differentiation
One of the earliest examples of regulated alternative splicing of pre-mRNA came from studies of sexual differentiation in Drosophila. The genes required for normal Drosophila sexual differentiation were first characterized by isolating Drosophila mutants defective in the process. When the proteins encoded by the wild-type genes were characterized biochemically, two of them were found to regulate a cascade of alternative RNA splicing in Drosophila embryos. More recent research has provided insight into how these proteins regulate RNA processing and ultimately lead to the creation of two different sex-specific transcriptional repressors that suppress the development of characteristics of the opposite sex.
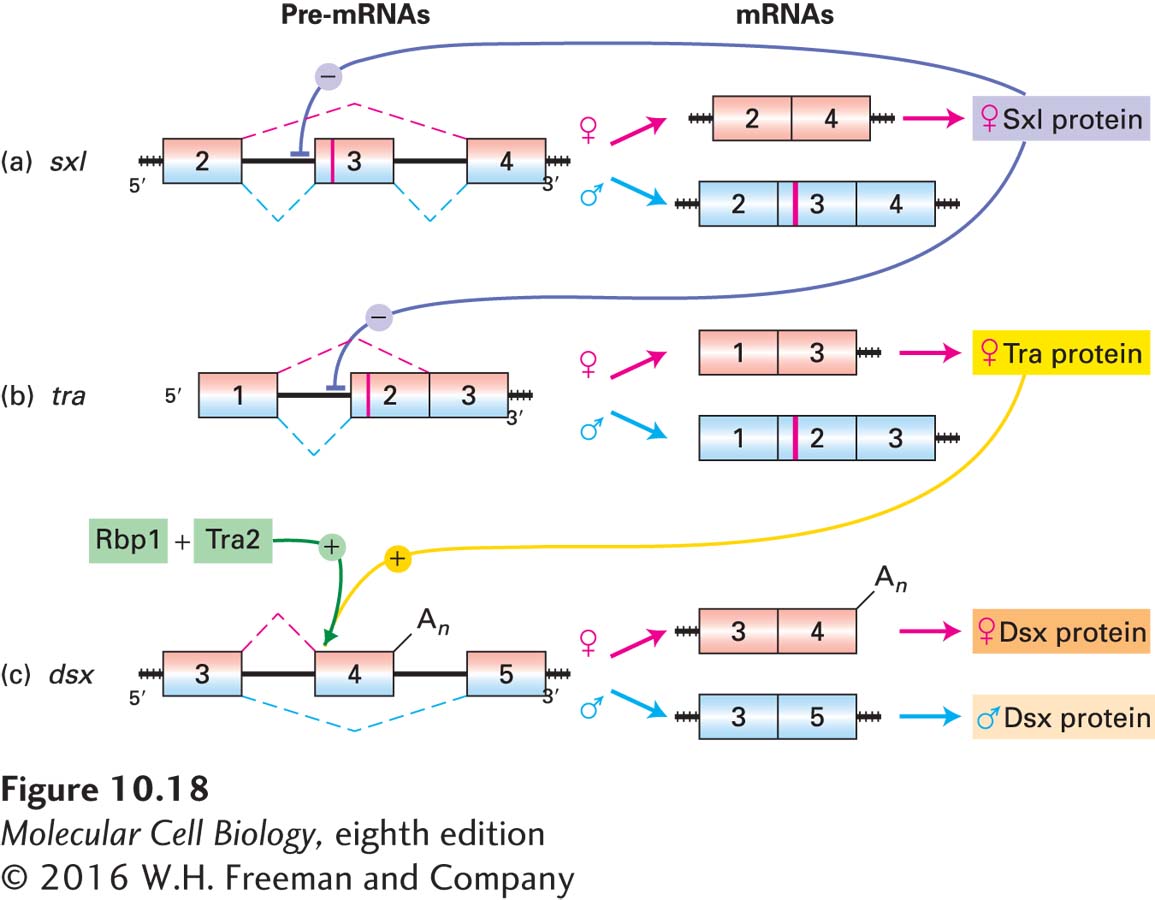
The Sex-lethal (Sxl) protein, encoded by the sex-lethal gene, is the first protein to act in the cascade (Figure 10-18). The Sxl protein is present only in female embryos. Early in development, the Sxl gene is transcribed from a promoter that functions only in female embryos. Later in development, this female-specific promoter is shut off, and another promoter for sex-lethal becomes active in both male and female embryos. In male embryos, however, in the absence of early Sxl protein, exon 2 of the sex-lethal pre-mRNA is spliced to exon 3 to produce an mRNA that contains a stop codon early in the sequence. The net result is that male embryos produce no functional Sxl protein either early or later in development.
FIGURE 10-18A cascade of regulated splicing controls sex determination in Drosophila embryos. For clarity, only the exons (boxes) and introns (black lines) where regulated splicing occurs are shown. Splicing is indicated by red dashed lines above (female) and blue dashed lines below (male) the pre-mRNAs. Vertical red lines in exons indicate in-frame stop codons, which prevent synthesis of functional protein. Only female embryos produce functional Sxl protein, which represses splicing between exons 2 and 3 in sxl pre-mRNA (a) and between exons 1 and 2 in tra pre-mRNA (b). (c) In contrast, the cooperative binding of Tra protein and two SR proteins, Rbp1 and Tra2, activates splicing between exons 3 and 4 and cleavage/polyadenylation(An) at the 3′ end of exon 4 in dsx pre-mRNA in female embryos. In male embryos, which lack functional Tra, the SR proteins do not bind to exon 4, and consequently exon 3 is spliced to exon 5. The distinct Dsx proteins produced in female and male embryos as the result of this cascade of regulated splicing repress transcription of genes required for sexual differentiation of the opposite sex. See M. J. Moore et al., 1993, in R. Gesteland and J. Atkins, eds., The RNA World, Cold Spring Harbor Press, pp. 303–357.
Page 436
In contrast, the Sxl protein expressed in early female embryos regulates splicing of the sex-lethal pre-mRNA so that a functional sex-lethal mRNA is produced (Figure 10-18a). Sxl accomplishes this by binding to a sequence in the pre-mRNA near the 3′ end of the intron between exon 2 and exon 3, thereby blocking the proper association of U2AF and U2 snRNP with the adjacent 3′ splice site used in males (see Figure 10-11). As a consequence, the U1 snRNP bound to the 5′ end of the intron between exons 2 and 3 assembles into a spliceosome with U2 snRNP bound to the branch point at the 3′ end of the intron between exons 3 and 4, leading to the splicing of exon 2 to exon 4 and the skipping of exon 3. The binding site for Sxl in the sex-lethal pre-mRNA is called an intronic splicing silencer because of its location in an intron and its function in blocking, or “silencing,” the use of a splice site. The resulting female-specific sex-lethal mRNA is translated into functional Sxl protein, which reinforces its own expression in female embryos by continuing to cause skipping of exon 3. The absence of Sxl protein in male embryos allows the inclusion of exon 3 and, consequently, of the stop codon near the 5′ end of exon 3 that prevents translation of functional Sxl protein (see Figure 10-18a).
Sxl protein also regulates alternative splicing of the pre-mRNA transcribed from the transformer gene (Figure 10-18b). In male embryos, in which no Sxl is expressed, exon 1 is spliced to exon 2, which contains a stop codon that prevents synthesis of a functional Transformer (Tra) protein. In female embryos, however, binding of Sxl protein to an intronic splicing silencer at the 3′ end of the intron between exons 1 and 2 blocks binding of U2AF at this site. The interaction of Sxl with transformer pre-mRNA is mediated by two adjacent RRM domains in the protein (see Figure 10-5). When Sxl is bound, U2AF binds to a lower-affinity site farther 3′ in the pre-mRNA; as a result, exon 1 is spliced to this alternative 3′ splice site, causing skipping of exon 2 with its stop codon. The resulting female-specific transformer mRNA, which contains additional constitutively spliced exons, is translated into functional Tra protein.
Finally, Tra protein regulates the alternative processing of pre-mRNA transcribed from the doublesex (dsx) gene (Figure 10-18c). In female embryos, a complex of Tra and two constitutively expressed SR proteins, Rbp1 and Tra2, directs the splicing of exon 3 to exon 4 and also promotes cleavage/polyadenylation at the alternative poly(A) site at the 3′ end of exon 4, leading to a short, female-specific version of the Dsx protein. In male embryos, which produce no Tra protein, exon 4 is skipped, so that exon 3 is spliced to exon 5. Exon 5 is constitutively spliced to exon 6, which is polyadenylated at its 3′ end—leading to a longer, male-specific version of the Dsx protein. The RNA sequence to which Tra binds in exon 4 is called an exonic splicing enhancer because it enhances splicing at a nearby splice site.
As a result of the cascade of regulated RNA processing depicted in Figure 10-18, different Dsx proteins are expressed in male and female embryos. The two proteins are transcription factors that share the N-terminal sequence encoded in exons 1–3, including a common DNA-binding domain, but have different C-terminal sequences, encoded by exon 4 in females and exon 5 plus additional downstream exons in males. The unique C-terminal end of the female protein functions as a strong activation domain, while the C-terminal end of the male protein is a strong repression domain. Consequently, the female Dsx protein activates genes with binding sites for the transcription factor, including genes that induce development of female characteristics, while the male Dsx protein represses the same target genes.
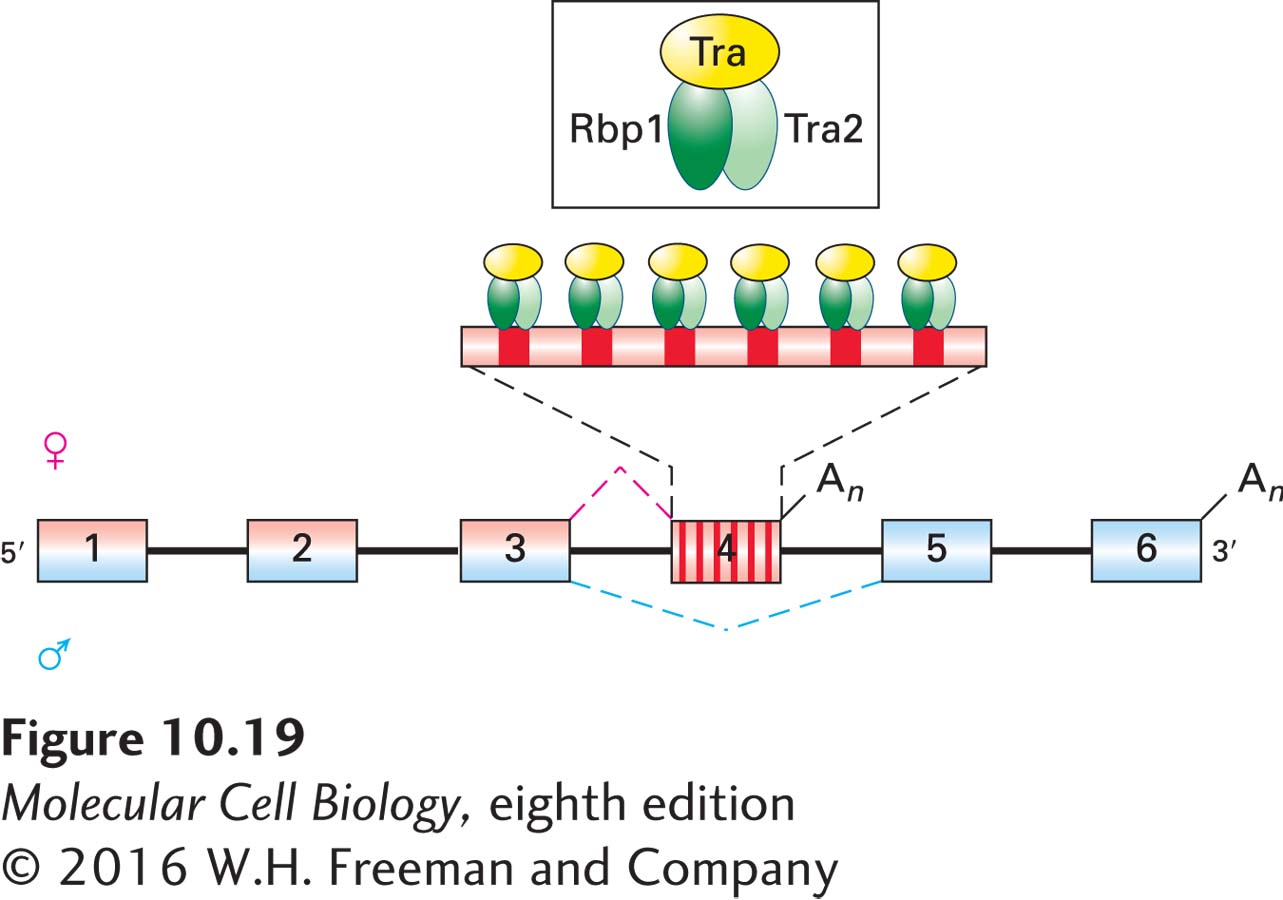
Figure 10-19 illustrates how the Tra/Tra2/Rbp1 complex is thought to interact with doublesex pre-mRNA. Rbp1 and Tra2 are SR proteins, but they do not interact with exon 4 in the absence of the Tra protein. The interaction of the Tra protein with Rbp1 and Tra2 results in the cooperative binding of all three proteins to six exonic splicing enhancers in exon 4. The bound Tra2 and Rbp1 proteins then promote the binding of U2AF and the U2 snRNP to the 3′ end of the intron between exons 3 and 4, just as other SR proteins do for constitutively spliced exons (see Figure 10-13). The Tra/Tra2/Rbp1 complexes also enhance binding of the cleavage/polyadenylation complex to the 3′ end of exon 4 because the U2 snRNP plus associated proteins bound to a 3′ splice site enhance binding of cleavage/polyadenylation factors (see Figure 10-15) to an appropriately spaced polyadenylation signal through cooperative binding interactions.
FIGURE 10-19Model of splicing activation by Tra protein and the SR proteins Rbp1 and Tra2. In female Drosophila embryos, splicing of exons 3 and 4 in dsx pre-mRNA is activated by the binding of Tra/Tra2/Rbp1 complexes to six exonic splicing enhancers in exon 4. Because Rbp1 and Tra2 cannot bind to the pre-mRNA in the absence of Tra, exon 4 is skipped in male embryos. See the text for discussion. An = polyadenylation. See T. Maniatis and B. Tasic, 2002, Nature418:236.