Surveillance Mechanisms Prevent Translation of Improperly Processed mRNAs
Translation of an improperly processed mRNA could lead to production of an abnormal protein that interferes with the gene’s normal function. This effect would be equivalent to that of a dominant-negative mutation, discussed in Chapter 6 (see Figure 6-41). Several mechanisms, collectively termed mRNA surveillance, help cells avoid the translation of improperly processed mRNA molecules. We have previously mentioned two such surveillance mechanisms: the recognition of improperly processed pre-mRNAs in the nucleus and their degradation by nuclear exosomes, and the general restriction against nuclear export of incompletely spliced pre-mRNAs that remain associated with a snRNP.
Another surveillance mechanism, called nonsense-mediated decay (NMD), causes degradation of mRNAs in which one or more exons have been incorrectly spliced. Such incorrect splicing often alters the open reading frame of the mRNA 3′ to the improper exon-exon junction, resulting in the introduction of an out-of-frame missense mutation and an incorrect stop codon. For nearly all properly spliced mRNAs, the stop codon is in the last exon. Nonsense-mediated decay results in the rapid degradation of mRNAs with stop codons that occur before the last exon-exon junction, since in most cases, such mRNAs arise from errors in RNA splicing. However, NMD can also result from a mutation creating a stop codon within a gene or a frame-shifting deletion or insertion. NMD was initially discovered during the study of patients with β0-thalassemia, who produce a low level of β-globin protein associated with a low level of β-globin mRNA (Figure 10-34).
[Part (b) republished with permission of Elsevier, from Maquat, L.E., et al., “Unstable β-globin mRNA in mRNA-deficient β0 thalassemia,” Cell, 1981, 27(3 Pt2):543–53; permission conveyed through Copyright Clearance Center, Inc.]
FIGURE 10-34Discovery of nonsense-mediated decay (NMD). (a) Patients with β0-thalassemia express very low levels of β-globin mRNA. A common cause of this syndrome is a single-base-pair deletion in exon 1 or exon 2 of the β-globin gene. Ribosomes translating the mutant mRNA read out of frame following the deletion and encounter a stop codon in the wrong reading frame before they translate across the last exon-exon junction in the mRNA. Consequently, they leave an exon-junction complex (EJC) in place on the mRNA. Cytoplasmic proteins associate with the EJC and induce degradation of the mRNA. (b) Bone marrow was obtained from a patient with a wild-type β-globin gene and from a patient with β0-thalassemia. RNA was isolated from the bone marrow cells shortly after collection and again 30 minutes after incubation in media with actinomycin D, a drug that inhibits transcription. The amount of β-globin RNA was measured using the S1-nuclease protection method (arrow). The patient with β0-thalassemia had much less β-globin mRNA than the patient with a wild-type β-globin gene (−Act D). The mutant β-globin mRNA decayed rapidly when transcription was inhibited (+Act D), whereas the wild-type β-globin mRNA remained stable.
[Part (b) republished with permission of Elsevier, from Maquat, L.E., et al., “Unstable β-globin mRNA in mRNA-deficient β0 thalassemia,” Cell, 1981, 27(3 Pt2):543–53; permission conveyed through Copyright Clearance Center, Inc.]
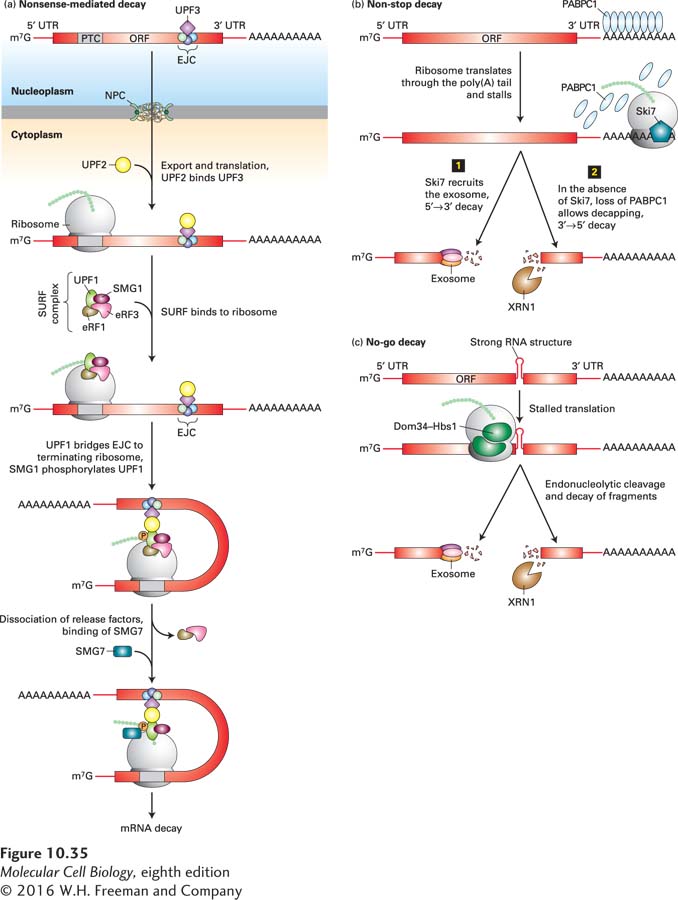
A search for possible molecular signals that might indicate the positions of exon-exon junctions in a processed mRNA led to the discovery of exon-junction complexes. As noted already, these complexes of several proteins (including Y14, Magoh, eIF4IIIA, UPF2, UPF3, and REF) bind about 20 nucleotides 5′ to an exon-exon junction following RNA splicing and stimulate export of mRNPs from the nucleus by interacting with the mRNP exporter (see Figure 10-23). Analysis of yeast mutants indicated that one of the proteins in exon-junction complexes (UPF3) functions in nonsense-mediated decay. In the cytoplasm, UPF3 interacts with a protein (UPF1) and a protein kinase (SMG1) that phosphorylates it, causing the mRNA to associate with P bodies, repressing translation and inducing degradation of the mRNA. An additional protein (UPF2) associated with the exon-junction complex binds a P body–associated deadenylase complex that rapidly removes the poly(A) tail from the associated mRNA, leading to its decapping and degradation by the P body–associated 5′→3′ exoribonuclease XRN1 (see Figure 10-24). In the case of a properly spliced mRNA, the mRNP exporter associated with the nuclear cap-binding complex is exported through a nuclear pore complex, thereby protecting the mRNA from degradation. The exon-junction complexes are thought to be dislodged from the mRNA by passage of the first “pioneer” ribosome to translate the mRNA. However, for mRNAs with a stop codon before the final exon-exon junction, one or more exon-junction complexes remain associated with the mRNA, resulting in nonsense-mediated decay (Figure 10-35a). Alternative mechanisms lead to the inhibition of translation and degradation of mRNAs that were polyadenylated prematurely (non-stop decay) (Figure 10-35b) or that contain damaged bases or stable secondary structures that block ribosomal translocation along the mRNA (no-go decay) (Figure 10-35c).
Page 457
FIGURE 10-35Mechanisms of RNA surveillance in the cytoplasm. (a) Nonsense-mediated decay. PTC = premature termination (stop) codon; SURF complex = complex of protein kinase SMG1, UPF1, and release factors eRF1 and eRF3. Formation of the SURF complex leads to phosphorylation of UPF1 by SMG1. The phosphorylated UPF1 associates with the UPF2-UPF3 complex bound to any exon-exon junction complexes that were not displaced from the mRNA by the first, pioneer ribosome to translate the message. This association leads to the association of the PTC-containing mRNA with P bodies, removal of the poly(A) tail, and degradation of the mRNA. (b) Non-stop decay. mRNAs that were prematurely cleaved and polyadenylated do not contain a stop codon before the poly(A) tail. When such mRNAs are translated, the ribosome translates the poly(A) tail and stalls at the 3′ end of the abnormal mRNA because the stop codon required for release factors eRF1 and eRF3 to associate with the ribosome A site is absent (see Figure 5-26). In higher eukaryotes, the factor Ski7 binds to the stalled ribosome and recruits the cytoplasmic exosome, which degrades the abnormal RNA step 1. Alternatively, in S. cerevisiae step 2, the displacement of PABPC1 from the poly(A) tail by the elongating ribosome leads to decapping and 5′→3′ degradation by the XRN1 exonuclease. (c) No-go decay. If a base of an mRNA is damaged so that a ribosome stalls there, or if an improperly processed mRNA has a stable stem-loop region with a long duplex stem that blocks elongation by the ribosome, the Dom34-Hbs1 complex binds the abnormal mRNA and makes an endonucleolytic cut in it. This cut generates a free 3′ end on the 5′ fragment, which is degraded by a cytoplasmic exosome, and a free 5′ end on the 3′ fragment, which is digested in the 5′→3′ direction by the XRN1 exonuclease. See N. L. Garneau, J. Wilusz, and C. J. Wilusz, 2007, Nat. Rev. Mol. Cell Biol.8:113.