Self-Splicing Group I Introns Were the First Examples of Catalytic RNA
During the 1970s, the pre-rRNA genes of the protozoan Tetrahymena thermophila were discovered to contain an intron. Careful searches failed to uncover even one pre-rRNA gene without the extra sequence, indicating that splicing is required to produce mature rRNAs in these organisms. In 1982, in vitro studies showing that the pre-rRNA is spliced at the correct sites in the absence of any protein provided the first indication that RNA can function as a catalyst, as enzymes do.
A whole raft of self-splicing sequences were subsequently found in pre-rRNAs from other single-celled organisms, in mitochondrial and chloroplast pre-rRNAs, in several pre-mRNAs from certain E. coli bacteriophages, and in some bacterial tRNA primary transcripts. The self-splicing sequences in all these precursors, referred to as group I introns, use guanosine as a cofactor and can fold by internal base pairing to juxtapose the two exons that must be joined. As discussed earlier, certain mitochondrial and chloroplast pre-mRNAs and tRNAs contain a second type of self-splicing intron, designated group II introns.
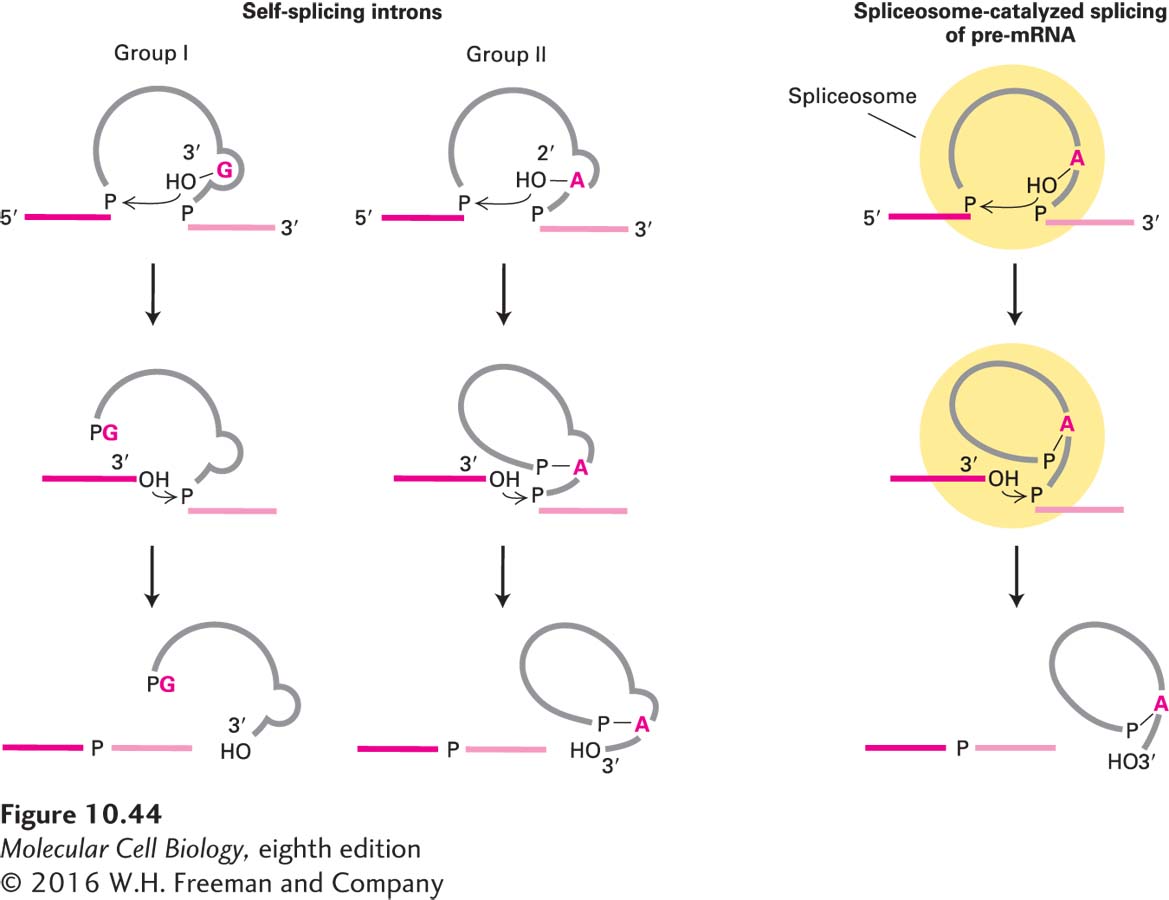
The splicing mechanisms used by group I introns, group II introns, and spliceosomes are generally similar, involving two transesterification reactions, which require no input of energy (Figure 10-44). Structural studies of the group I intron from Tetrahymena pre-rRNA, combined with mutational and biochemical experiments, have revealed that the RNA folds into a precise three-dimensional structure that, like an enzyme, contains deep grooves for binding substrates and solvent-inaccessible regions that function in catalysis. The group I intron functions like a metalloenzyme to precisely place the atoms that participate in the two transesterification reactions adjacent to catalytic Mg2+ ions. Considerable evidence now indicates that splicing by group II introns and by snRNAs in the spliceosome also involves bound catalytic Mg2+ ions. In both group I and group II self-splicing introns, and probably in the spliceosome, RNA functions as a ribozyme, an RNA sequence with catalytic ability.
FIGURE 10-44Splicing mechanisms in group I and group II self-splicing introns and in spliceosome-catalyzed splicing of pre-mRNA. The intron is shown in gray, the exons to be joined in red. In group I introns, a guanosine cofactor (G) that is not part of the RNA chain associates with the active site. The 3′-hydroxyl group of this guanosine participates in a transesterification reaction with the phosphate at the 5′ end of the intron; this reaction is analogous to that involving the 2′-hydroxyl groups of the branch-point As in group II introns and pre-mRNA introns spliced in spliceosomes (see Figure 10-8). The subsequent transesterification that links the 5′ and 3′ exons is similar in all three splicing mechanisms. Note that spliced-out group I introns are linear structures, unlike the branched intron products in the other two cases. See P. A. Sharp, 1987, Science235:769.