Splicing requires the presence of small nuclear RNAs (snRNAs), which base-pair with the pre-mRNA, and some 170 associated proteins. Five U-rich snRNAs, designated U1, U2, U4, U5, and U6, participate in pre-mRNA splicing. Ranging in length from 107 to 210 nucleotides, these snRNAs are associated with 6–10 proteins each in the many small nuclear ribonucleoprotein particles (snRNPs) in the nuclei of eukaryotic cells.
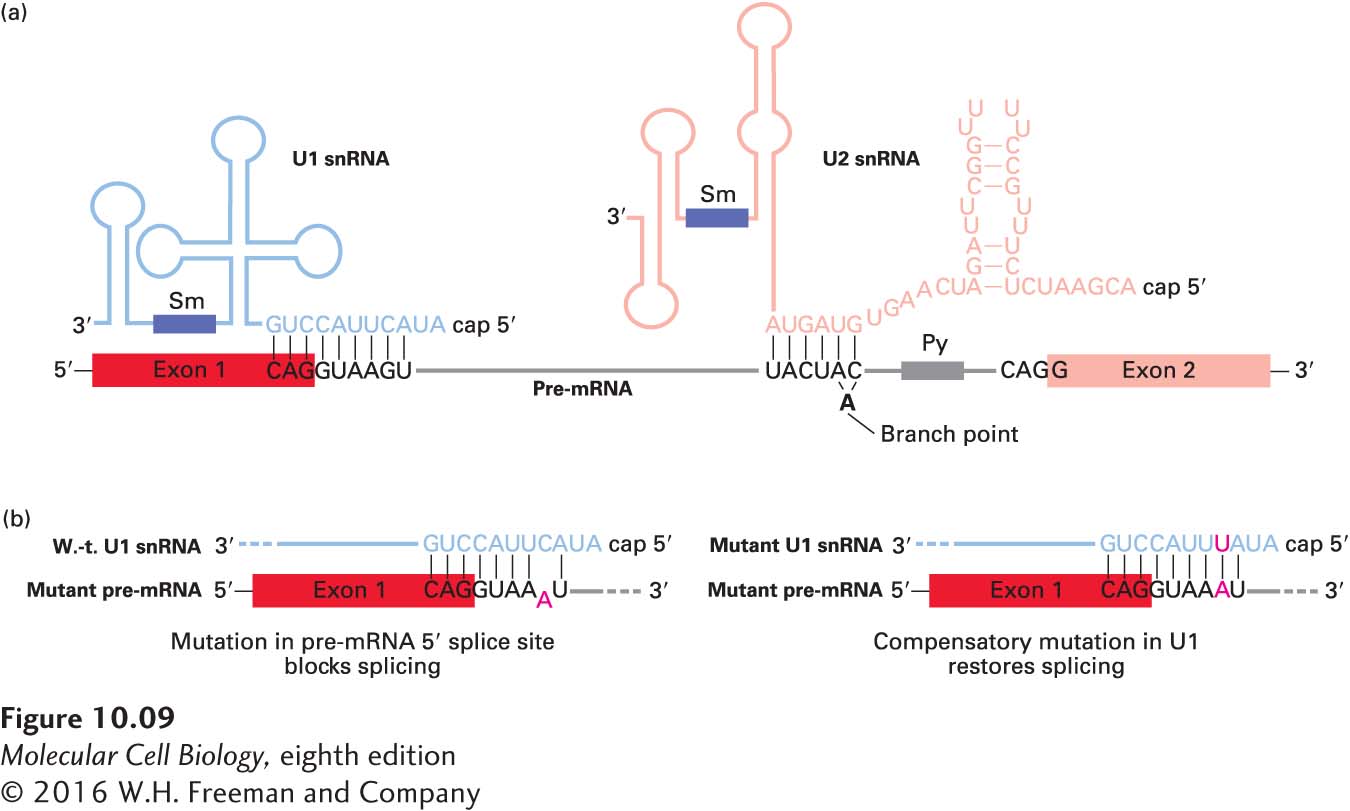
Definitive evidence for the role of U1 snRNA in splicing came from experiments indicating that base pairing between the 5′ splice site of a pre-mRNA and the 5′ region of U1 snRNA is required for RNA splicing (Figure 10-9a). In vitro experiments showed that a synthetic oligonucleotide that hybridizes with the 5′-end region of U1 snRNA blocks RNA splicing. In vivo experiments showed that base pairing–disrupting mutations in the pre-mRNA 5′ splice site also block RNA splicing; in this case, however, splicing can be restored by expression of a U1 snRNA with a compensating mutation that restores base pairing to the mutant pre-mRNA 5′ splice site (Figure 10-9b). Involvement of U2 snRNA in splicing was initially suspected when it was found to have an internal sequence that is largely complementary to the consensus sequence flanking the branch point in pre-mRNAs (see Figure 10-7). Compensating mutation experiments, similar to those conducted with U1 snRNA and 5′ splice sites, demonstrated that base pairing between U2 snRNA and the branch-point sequence in pre-mRNA is also critical to splicing.
FIGURE 10-9 Base pairing between pre-mRNA, U1 snRNA, and U2 snRNA early in the splicing process. (a) In this diagram, secondary structures in the snRNAs that are not altered during splicing are depicted schematically. The yeast branch-point sequence is shown here. Note that U2 snRNA base-pairs with a sequence that includes the branch-point A, although this residue is not base-paired. For unknown reasons, antisera from patients with the autoimmune disease systemic lupus erythematosus (SLE) contain antibodies to snRNP proteins, which have been useful in characterizing components of the splicing reaction; the purple rectangles represent sequences that bind snRNP proteins recognized by these anti-Sm antibodies. (b) Only the 5′ ends of U1 snRNAs and 5′ splice sites in pre-mRNAs are shown. (Left) A mutation (A) in a pre-mRNA splice site that interferes with base pairing to the 5′ end of U1 snRNA blocks splicing. (Right) Expression of a U1 snRNA with a compensating mutation (U) that restores base pairing also restores splicing of the mutant pre-mRNA. See M. J. Moore et al., 1993, in R. Gesteland and J. Atkins, eds., The RNA World, Cold Spring Harbor Press, pp. 303–357; see also Y. Zhuang and A. M. Weiner, 1986, Cell46:827.
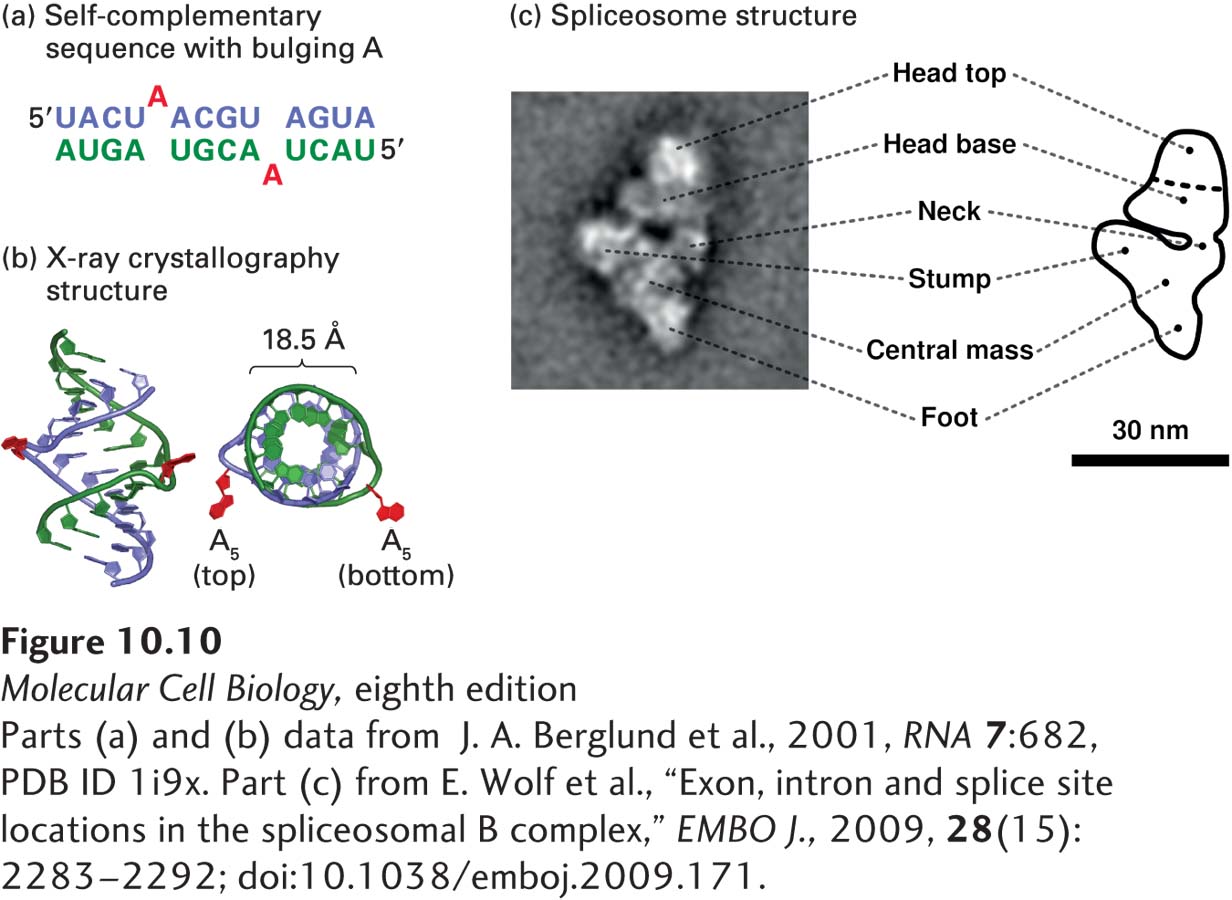
Figure 10-9a illustrates the general structures of the U1 and U2 snRNAs and how they base-pair with pre-mRNA during splicing. Significantly, the branch-point A itself, which is not base-paired to U2 snRNA, “bulges out” (Figure 10-10a), which allows its 2′ hydroxyl to participate in the first transesterification reaction of RNA splicing (see Figure 10-8).
Page 425
[Parts (a) and (b) data from J. A. Berglund et al., 2001, RNA7:682, PDB ID 1i9x. Part (c) from E. Wolf et al., “Exon, intron and splice site locations in the spliceosomal B complex,” EMBO J., 2009, 28(15):2283–2292; doi:10.1038/emboj.2009.171.]
FIGURE 10-10Structures of a bulged A in an RNA-RNA helix and an intermediate in the splicing process. (a) Diagram of RNA duplex used for determining the structure of a bulged A. Bulged As at position 5 (red) are excluded from duplex RNA-RNA hybrid formed by complementary bases (blue and green). (b) X-ray crystallography of the structure showed that the bulged A residues extend from the side of an A-form RNA-RNA helix. The phosphate backbone of one strand is shown in green and that of the other strand in blue. The structure on the right is turned 90 degrees for a view down the axis of the helix. (c) 40 Å resolution structure of a spliceosomal splicing intermediate containing U2, U4, U5, and U6 snRNPs, determined by cryoelectron microscopy and image reconstruction. The U4/U6/U5 tri-snRNP complex has a structure similar to the triangular body of this complex below the neck, suggesting that these snRNPs are at the bottom of the structure shown here and that the head is composed largely of U2 snRNP. See H. Stark and R. Luhrmann, 2006, Annu. Rev. Biophys. Biomol. Struct.35:435.
[Parts (a) and (b) data from J. A. Berglund et al., 2001, RNA7:682, PDB ID 1i9x. Part (c) from E. Wolf et al., “Exon, intron and splice site locations in the spliceosomal B complex,” EMBO J., 2009, 28(15):2283–2292; doi:10.1038/emboj.2009.171.]
Page 426
Similar studies with other snRNAs demonstrated that base pairing between the snRNAs themselves also occurs during splicing. Moreover, rearrangements in these RNA-RNA interactions are critical in the splicing pathway.
As mentioned above, a synthetic oligonucleotide that base pairs with the 5′ end of U1 snRNA was found to inhibit RNA splicing in vitro, supporting the importance of U1 snRNA base pairing to a 5′ splice site for the first step in pre-mRNA splicing. Currently, a similar strategy is being used in clinical trials for the treatment of Duchenne muscular dystrophy (DMD). This disorder is the most common human genetic disease due to new mutations in the genome. It is caused by mutations in the DMD gene, especially chain-terminating mutations due to a base-pair change in an exon that generates a stop codon. Alternatively, short deletions or insertions that change the reading frame of the message result in translation of abnormal amino acids, generally followed by a stop codon in the altered reading frame. These mutations eliminate the C-terminus of the encoded protein, dystrophin, which is essential to its function (see Figure 17-20, bottom). The DMD gene is the longest human gene (~2 million base pairs; half the length of the entire E. coli genome!), which makes it a large target for random mutations. Since the DMD gene is on the X chromosome, there is no second wild-type copy to complement the mutation in males. Synthetic oligonucleotides have been developed that are modified to permeate cell membranes, but have normal Watson-Crick base-pairing properties. By hybridizing with the terminus of a mutant exon, they can cause the abnormal exon to be “skipped” during pre-mRNA splicing, and can be designed so that the normal exon upstream of the mutation splices to an in-frame downstream exon. This results in expression of a protein with an internal deletion, but one that, potentially, has sufficient function to alleviate what are otherwise devastating symptoms.