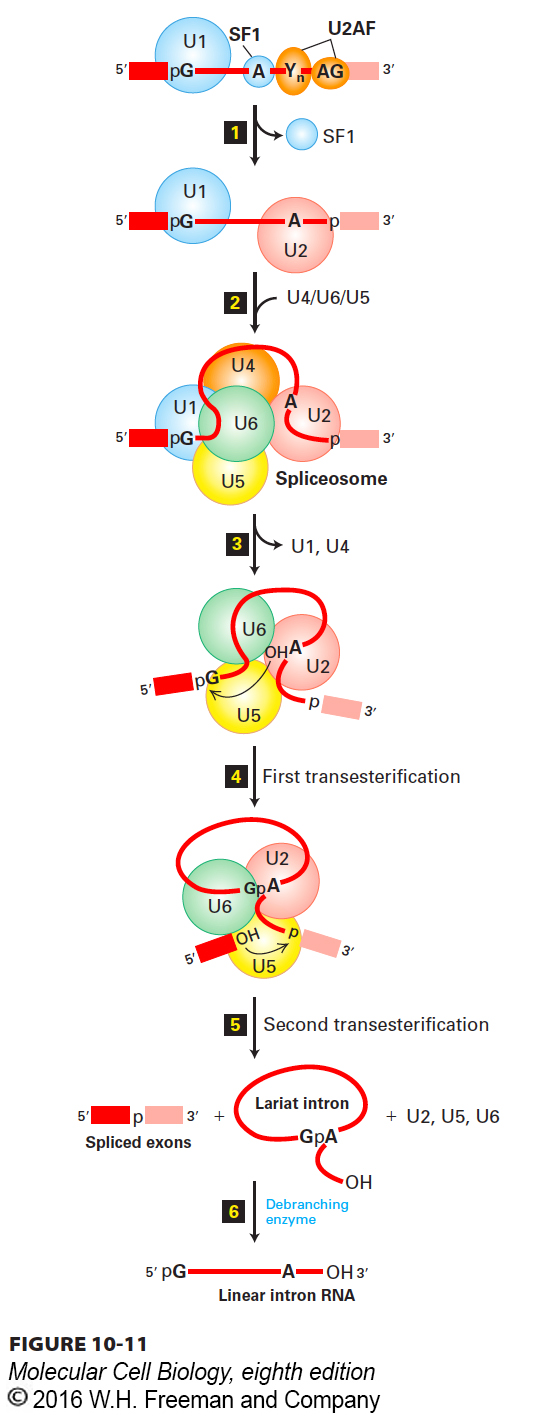
Spliceosomes, Assembled from snRNPs and a Pre-mRNA, Carry Out Splicing
FIGURE 10-11Model of spliceosome-mediated splicing of pre-mRNA. Step 1: After U1 base-pairs with the consensus 5′ splice site, SF1 (splicing factor 1) binds the branch-point A; U2AF (U2 snRNP associated factor) associates with the polypyrimidine tract and 3′ splice site; and the U2 snRNP associates with the branch-point A via base-pairing interactions shown in Figure 10-9, displacing SF1. Step 2: A trimeric snRNP complex of U4, U5, and U6 joins the initial complex to form the spliceosome. Step 3: Rearrangements of base-pairing interactions between snRNAs convert the spliceosome into a catalytically active conformation and destabilize the U1 and U4 snRNPs, which are released. Step 4: The catalytic core, thought to be formed by U6 and U2, then catalyzes the first transesterification reaction, forming the intermediate containing a 2′,5′-phosphodiester bond, as shown in Figure 10-8. Step 5: Following further rearrangements between the snRNPs, the second transesterification reaction joins the two exons by a standard 3′,5′-phosphodiester bond and releases the intron as a lariat structure as well as the remaining snRNPs. Step 6: The excised lariat intron is converted into a linear RNA by a debranching enzyme. See T. Villa et al., 2002, Cell109:149.
The five splicing snRNPs and other proteins involved in splicing assemble on a pre-mRNA, forming a large ribonucleoprotein complex called a spliceosome (Figure 10-11). The spliceosome has a mass similar to that of a ribosome. Assembly of a spliceosome begins with the base pairing of the U1 snRNA to the 5′ splice site as well as the cooperative binding of protein SF1 (splicing factor 1) to the branch-point A and of the heterodimeric protein U2AF (U2-associated factor) to the polypyrimidine tract and the 3′ AG of the intron via its large and small subunits, respectively. The U2 snRNP then base-pairs with the branch-point region (see Figure 10-9a) as SF1 is released. Extensive base pairing between the snRNAs in the U4 and U6 snRNPs forms a complex that associates with U5 snRNP. This U4/U6/U5 “tri-snRNP” then associates with the previously formed U1/U2/pre-mRNA complex to generate a spliceosome.
After formation of the spliceosome, extensive rearrangements in the pairing of snRNAs and the pre-mRNA lead to the release of the U1 snRNP. Figure 10-10c shows the structure of this intermediate in the splicing process. A further rearrangement of spliceosomal components occurs with the loss of the U4 snRNP. Its release generates a complex that catalyzes the first transesterification reaction that forms the 2′,5′-phosphodiester bond between the 2′ hydroxyl on the branch-point A and the phosphate at the 5′ end of the intron (see Figure 10-8). Following another rearrangement of the snRNPs, the second transesterification reaction ligates the two exons in a standard 3′,5′-phosphodiester bond, releasing the intron as a lariat structure associated with the snRNPs. This final intron-snRNP complex rapidly dissociates, and the individual snRNPs released can participate in a new cycle of splicing. The excised intron is then rapidly degraded by a debranching enzyme and other nuclear RNases discussed later.
Page 427
As mentioned above, a spliceosome is roughly the size of a ribosome and is composed of about 170 proteins, including about 100 “splicing factors” in addition to the proteins associated with the five snRNPs. This makes RNA splicing comparable in complexity to initiation of transcription and protein synthesis. Some of the splicing factors are associated with snRNPs, but others are not. For instance, the 65-kDa subunit of U2AF binds to the polypyrimidine tract near the 3′ end of an intron and to the U2 snRNP. The 35-kDa subunit of U2AF binds to the AG dinucleotide at the 3′ end of the intron and also interacts with the larger U2AF subunit bound nearby. These two U2AF subunits act together with SF1 to help specify the 3′ splice site by promoting interaction of the U2 snRNP with the branch point (see Figure 10-11, step 1). Some splicing factors also exhibit sequence homologies to known RNA helicases; these factors are probably necessary for the base-pairing rearrangements that occur among snRNAs during the spliceosomal splicing cycle. Several splicing factors associate with the CTD of RNA polymerase II when it is phosphorylated at serine 2 of the heptapeptide repeat by the cyclin T–CDK9 transcription elongation factor (see Figure 9-21). This association concentrates these splicing factors near the RNA exit site of RNA polymerase II so that they can rapidly assemble a spliceosome at a splice site as it emerges from the polymerase.
Following RNA splicing, a specific set of hnRNP proteins remains bound to the spliced RNA approximately 20 nucleotides 5′ to each exon-exon junction, thus forming an exon-junction complex. One of the hnRNP proteins associated with the exon-junction complex is the RNA export factor (REF), which functions in the export of fully processed mRNPs from the nucleus to the cytoplasm, as discussed in Section 10.3. Other proteins associated with the exon-junction complex function in a quality-control mechanism in the cytoplasm that leads to the degradation of improperly spliced mRNAs, known as nonsense-mediated decay (see Section 10.4).
A small fraction of pre-mRNAs (~1 percent in humans) contain introns whose splice sites do not conform to the standard consensus sequence. This class of introns begins with AU and ends with AC rather than following the usual “GU-AG rule” (see Figure 10-7). Splicing of this special class of introns occurs via a splicing cycle analogous to that shown in Figure 10-11, except that four novel, low-abundance snRNPs, together with the standard U5 snRNP, are involved.
Page 428
Nearly all functional mRNAs in vertebrate, insect, and plant cells are derived from a single molecule of the corresponding pre-mRNA by removal of internal introns and splicing of exons. However, in two types of protozoans—trypanosomes and euglenoids—mRNAs are constructed by splicing together separate RNA molecules. This process, referred to as trans-splicing, is also used in the synthesis of 10–15 percent of the mRNAs in the nematode (roundworm) Caenorhabditis elegans, an important model organism for studying embryonic development. Trans-splicing is carried out by snRNPs by a process similar to the splicing of exons in a single pre-mRNA.