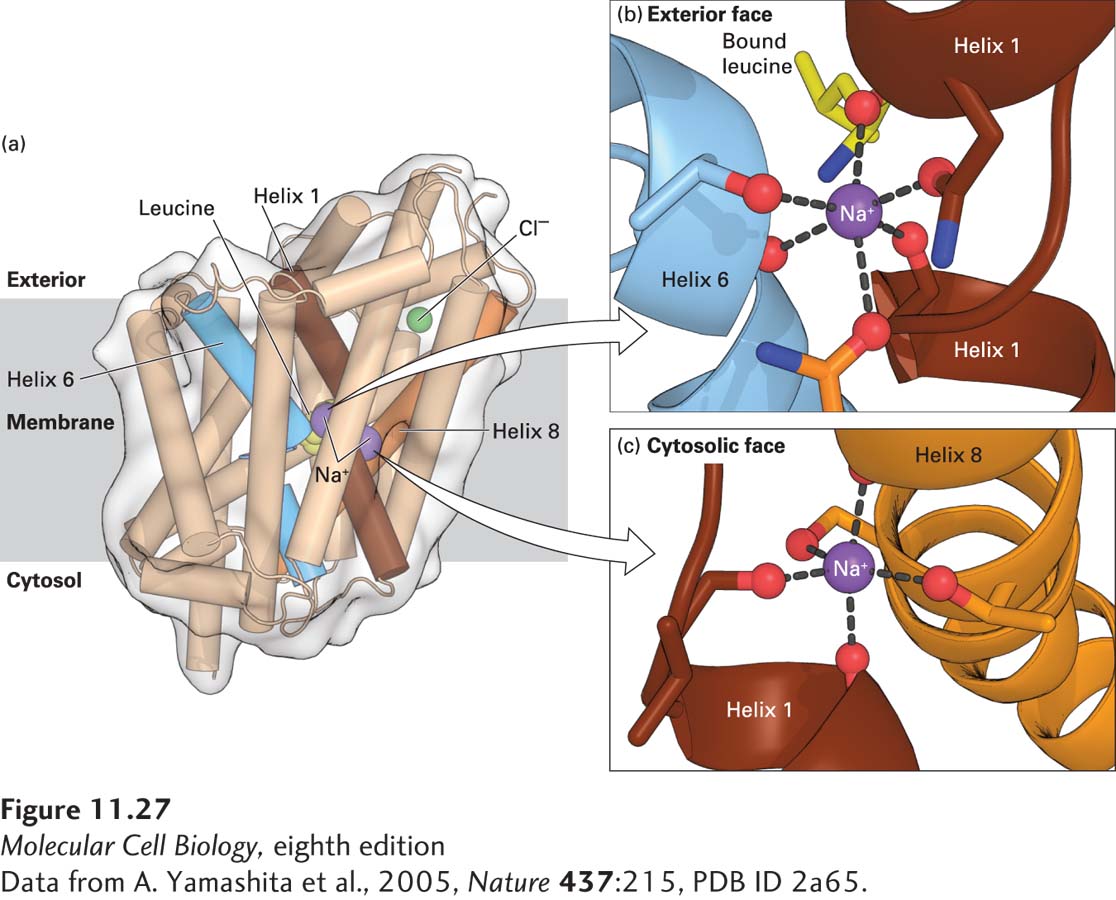
A Bacterial Na+/Amino Acid Symporter Reveals How Symport Works
No three-dimensional structure has yet been determined for any mammalian Na+-linked symporter, but the structures of several homologous bacterial Na+/substrate symporters have provided considerable information about symporter function. The bacterial two-Na+/one-leucine symporter shown in Figure 11-27a consists of 12 membrane-spanning α helices. Two of the helices (numbers 1 and 6) have nonhelical segments in the middle of the membrane that form part of the leucine-binding site.
[Data from A. Yamashita et al., 2005, Nature437:215, PDB ID 2a65.]
FIGURE 11-27Three-dimensional structure of the two-Na+/one-leucine symporter from the bacterium Aquifex aeolicus. (a) The bound L-leucine, two Na+ ions, and a Cl− ion are shown in yellow, purple, and green, respectively. The three membrane-spanning α helices that bind the Na+ or the leucine are colored brown, blue, and orange. (b, c) Binding of the two Na+ ions to carbonyl main-chain or carboxyl side-chain oxygen atoms (red) that are part of helices 1 (brown), 6 (blue), or 8 (orange). It is important that one of the Na+ ions is also bound to the carboxyl group of the transported leucine (part b). See H. Krishnamurthy et al., 2009, Nature459:347–355 for details on the structure and function of this and related Na+-linked symporters.
[Data from A. Yamashita et al., 2005, Nature437:215, PDB ID 2a65.]
The amino acid residues involved in binding the leucine and the two Na+ ions are located in the middle of the membrane-spanning segment (as depicted for the two-Na+/one-glucose symporter in Figure 11-26) and are close together in three-dimensional space. This proximity suggests that the coupling of amino acid and ion transport in these transporters is the consequence of direct or nearly direct physical interactions of the substrates. Indeed, one of the Na+ ions is bound to the carboxyl group of the transported leucine (Figure 11-27b). Thus neither substance can bind to the transporter without the other. Each of the two Na+ ions is bound to six oxygen atoms in the transporter. Sodium 1, for example, is bound to carbonyl oxygens of several transporter amino acids as well as to carbonyl oxygens and the hydroxyl oxygen of one threonine. Equally importantly, there are no water molecules surrounding either of the bound Na+ atoms, as is the case for K+ ions in potassium channels (see Figure 11-21). Thus as the Na+ ions lose their water of hydration in binding to the transporter, they bind to six oxygen atoms with a similar geometry. This reduces the energy change required for the binding of Na+ ions and prevents other ions, such as K+, from binding in place of Na+.
One striking feature of the structure depicted in Figure 11-27 is that the bound Na+ ions and leucine are occluded—that is, they cannot diffuse out of the protein to either the surrounding extracellular or cytoplasmic media. This structure represents an intermediate in the transport process (see Figure 11-26) in which the transporter appears to be changing from a conformation with an exoplasmic-facing to one with a cytosolic-facing binding site.