Numerous Transport Proteins Enable Plant Vacuoles to Accumulate Metabolites and Ions
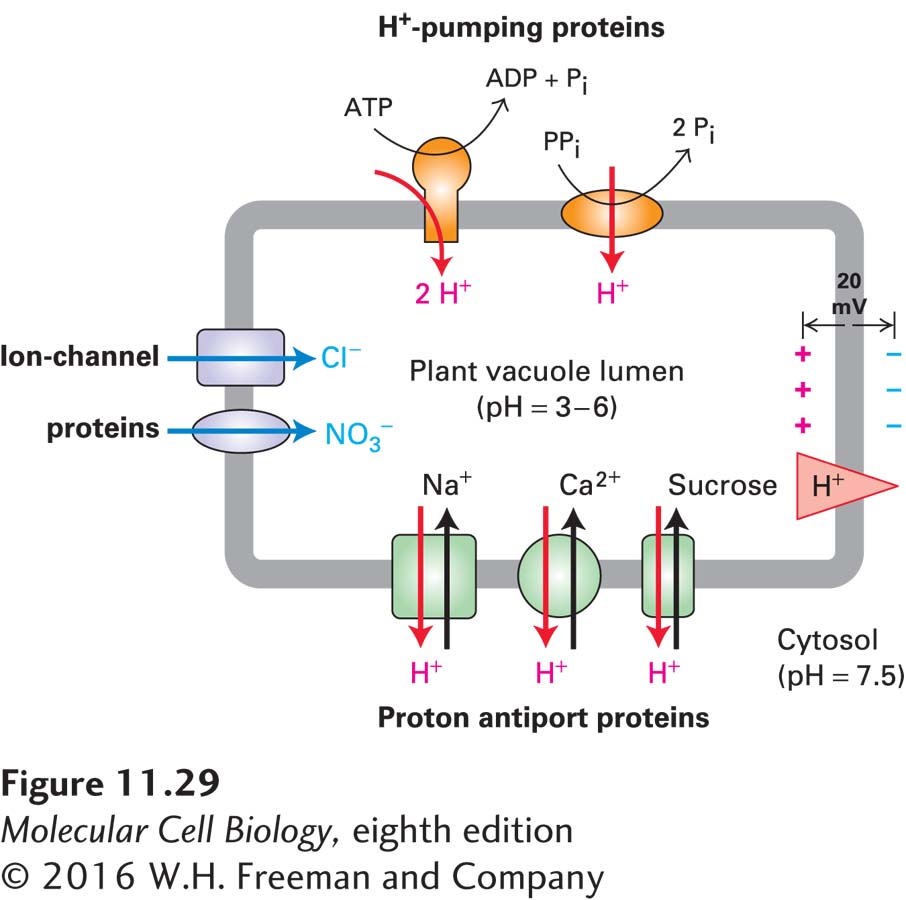
The lumen of a plant vacuole is much more acidic (pH 3–6) than is the cytosol (pH 7.5). The acidity of vacuoles is maintained by a V-class ATP-powered proton pump (see Figure 11-9) and by a pyrophosphate-powered proton pump that is unique to plants. Both of these pumps, located in the vacuolar membrane, import H+ ions into the vacuolar lumen against a concentration gradient. The vacuolar membrane also contains Cl− and NO3− channels that transport these anions from the cytosol into the vacuole. Entry of these anions against their concentration gradients is driven by the inside-positive electric potential generated by the proton pumps. The combined operation of these proton pumps and anion channels produces an inside-positive electric potential of about 20 mV across the vacuolar membrane as well as a substantial pH gradient (Figure 11-29).
FIGURE 11-29Concentration of ions and sucrose by the plant vacuole. The vacuolar membrane contains two types of proton pumps (orange): a V-class H+ ATPase (left) and a pyrophosphate-hydrolyzing proton pump (right) that differs from all other ion pumps and is probably unique to plants. These pumps generate a low luminal pH as well as an inside-positive electric potential across the vacuolar membrane by their inward pumping of H+ ions. The inside-positive potential powers the movement of Cl− and NO3− from the cytosol through separate channel proteins (purple). Proton antiporters (green), powered by the H+ gradient, accumulate Na+, Ca2+, and sucrose inside the vacuole. See B. J. Barkla and O. Pantoja, 1996, Annu. Rev. Plant Phys.47:159–184 and P. A. Rea et al., 1992, Trends Biochem. Sci.17:348.
The proton electrochemical gradient across the plant vacuole membrane is used in much the same way as the Na+ electrochemical gradient across the animal-cell plasma membrane: to power the selective uptake or extrusion of ions and small molecules by various antiporters. In the leaf, for example, excess sucrose generated by photosynthesis during the day is stored in the vacuole; during the night, the stored sucrose moves into the cytoplasm and is metabolized to CO2 and H2O with concomitant generation of ATP from ADP and Pi. A proton/sucrose antiporter in the vacuolar membrane operates to accumulate sucrose in plant vacuoles. The inward movement of sucrose is powered by the outward movement of H+, which is favored by its concentration gradient (lumen > cytosol) and by the cytosolic-negative potential across the vacuolar membrane (see Figure 11-29). Uptake of Ca2+ and Na+ into the vacuole from the cytosol against their concentration gradients is similarly mediated by proton antiporters.
Our understanding of the transporters in plant vacuolar membranes has the potential for increasing agricultural production in soils with a high salt (NaCl) concentration, which are found throughout the world. Because most agriculturally useful crops cannot grow in such saline soils, agricultural scientists have long sought to develop salt-tolerant plants by traditional breeding methods. With the availability of the cloned gene encoding the vacuolar Na+/H+ antiporter, researchers can now produce transgenic plants that overexpress this transporter, leading to increased sequestration of Na+ in the vacuole. For instance, transgenic tomato plants that overexpress the vacuolar Na+/H+ antiporter can grow, flower, and produce fruit in the presence of soil NaCl concentrations that kill wild-type plants. Interestingly, although the leaves of these transgenic tomato plants accumulate large amounts of salt, the fruit has a very low salt content.