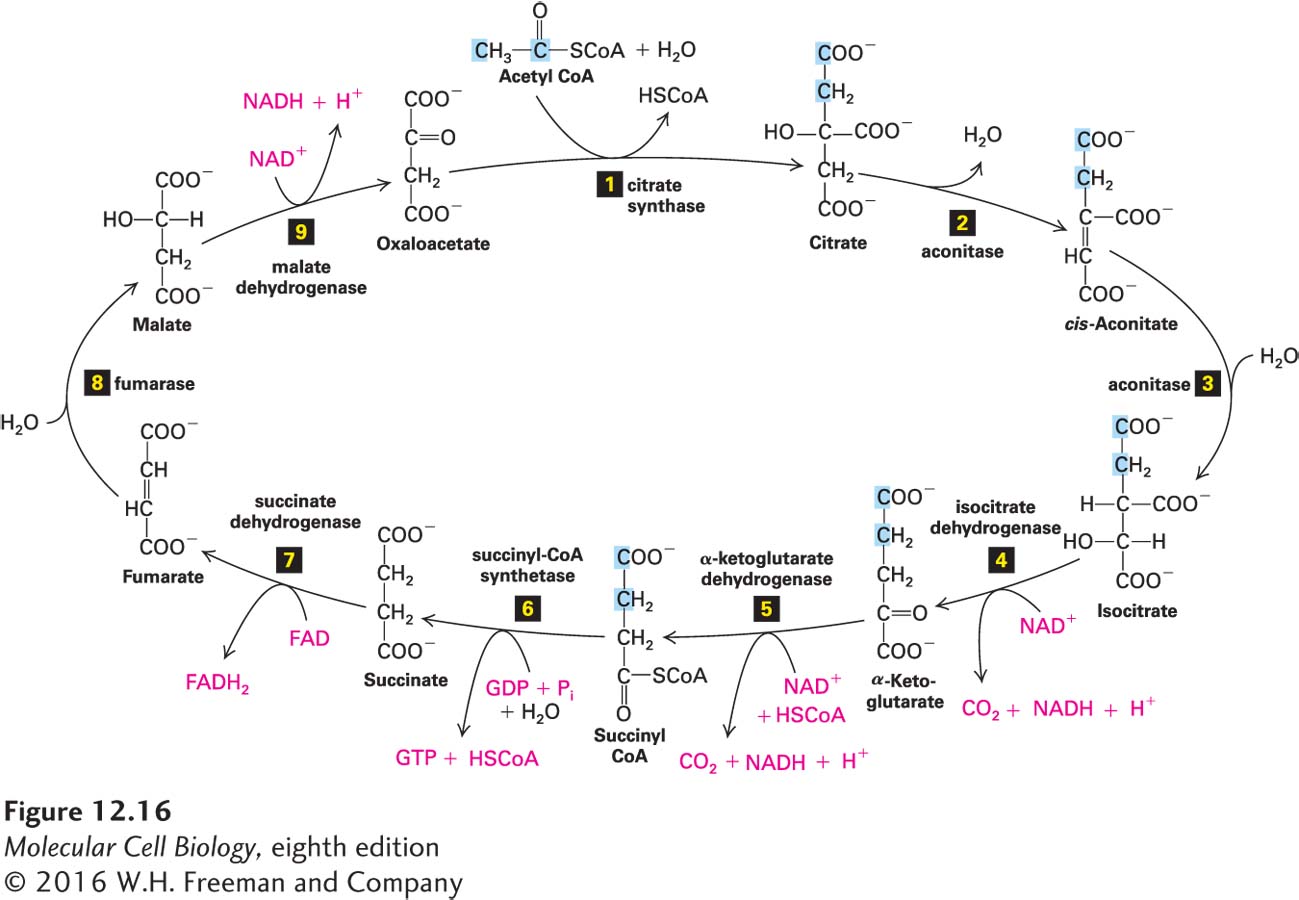
In the Second Part of Stage II, the Citric Acid Cycle Oxidizes the Acetyl Group in Acetyl CoA to CO2 and Generates High-Energy Electrons
Nine sequential reactions operate in a cycle to oxidize the acetyl group of acetyl CoA to CO2 (Figure 12-14, stage II, right). This cycle is referred to by several names: the citric acid cycle, the tricarboxylic acid (TCA) cycle, and the Krebs cycle. The net result is that for each acetyl group entering the cycle as acetyl CoA, two molecules of CO2, three of NADH, and one each of FADH2 and GTP are produced. NADH and FADH2 are high-energy electron carriers that will play a major role in stage III of mitochondrial oxidation: electron transport.
As shown in Figure 12-16, the cycle begins with condensation of the two-carbon acetyl group from acetyl CoA and the four-carbon molecule oxaloacetate to yield the six-carbon citric acid (hence the name citric acid cycle). Reactions step 4 and step 5 each release a CO2 molecule and reduce NAD+ to NADH. The source of the oxygen for generating the CO2 molecules in these reactions is water (H2O), not molecular oxygen (O2), and the enzymatic activities of the enzymes catalyzing reactions step 4 and step 5 are increased by the influx of calcium into the mitochondrion from the MAM. Reduction of NAD+ to NADH also occurs during reaction step 9; thus three NADHs are generated per turn of the cycle. In reaction step 7, two electrons and two protons are transferred to FAD, yielding the reduced form of this coenzyme, FADH2. Reaction step 7 is distinctive not only because it is an intrinsic part of the citric acid cycle (stage II), but also because it is catalyzed by a membrane-attached enzyme that, as we shall see, also plays an important role in stage III. In reaction step 6, hydrolysis of the high-energy thioester bond in succinyl CoA is coupled to synthesis of one GTP by substrate-level phosphorylation. Because GTP and ATP are interconvertible,
GTP + ADP ⇌ GDP + ATP
this can be considered an ATP-generating step. Reaction step 9 regenerates oxaloacetate, so the cycle can begin again. Note that molecular O2 does not participate in the citric acid cycle.
FIGURE 12-16The citric acid cycle. Acetyl CoA is metabolized to CO2 and the high-energy electron carriers NADH and FADH2. In reaction 1, a two-carbon acetyl residue from acetyl CoA condenses with the four-carbon molecule oxaloacetate to form the six-carbon citrate. In the remaining reactions (2–9), each molecule of citrate is eventually converted back to oxaloacetate, losing two CO2 molecules in the process. In each turn of the cycle, four pairs of electrons are removed from carbon atoms, forming three molecules of NADH, one molecule of FADH2, and one molecule of GTP. The two carbon atoms that enter the cycle with acetyl CoA are highlighted in blue through succinyl CoA. In succinate and fumarate, which are symmetric molecules, they can no longer be specifically denoted. Isotope-labeling studies have shown that these carbon atoms are not lost in the turn of the cycle in which they enter; on average, one will be lost as CO2 during the next turn of the cycle and the other in subsequent turns.
Page 534
Most enzymes and small molecules involved in the citric acid cycle are soluble in the aqueous mitochondrial matrix. These include CoA, acetyl CoA, succinyl CoA, NAD+, and NADH, as well as most of the citric acid cycle enzymes. Succinate dehydrogenase (reaction step 7), however, is a component of an integral membrane protein in the inner membrane, with its active site facing the matrix. When mitochondria are disrupted by gentle ultrasonic vibration or by osmotic lysis, the non-membrane-bound enzymes of the citric acid cycle are released as very large multiprotein complexes. It is believed that within such complexes, the reaction product of one enzyme passes directly to the next enzyme without diffusing through the solution (see Figure 3-30).
Page 535
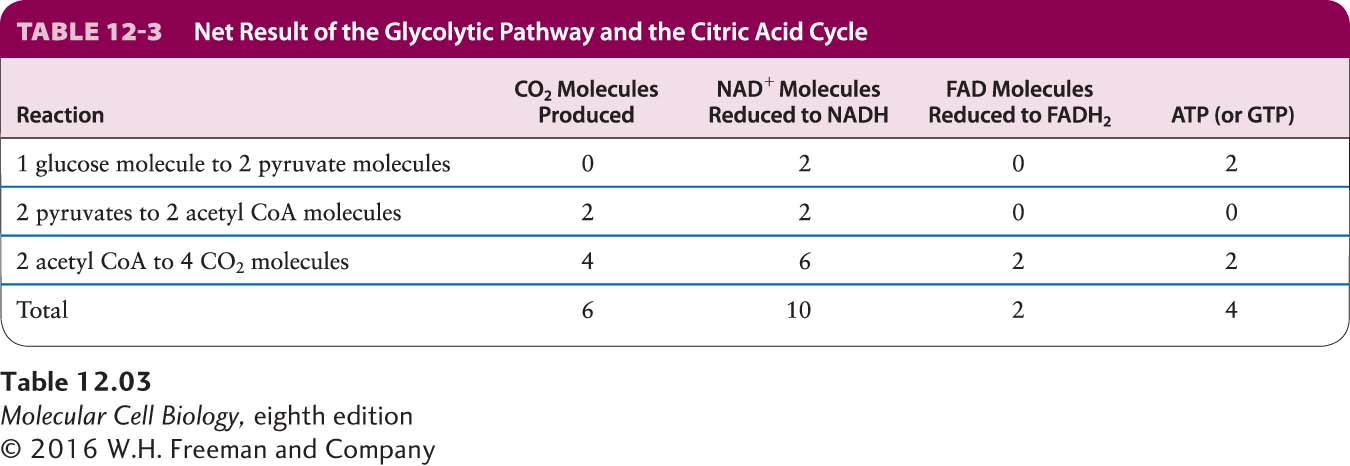
Because glycolysis of one glucose molecule generates two pyruvate molecules, and thus two acetyl CoA molecules, the reactions in the glycolytic pathway and citric acid cycle produce six CO2 molecules, ten NADH molecules, and two FADH2 molecules per glucose molecule (Table 12-3). Although these reactions also generate four high-energy phosphoanhydride bonds in the form of two ATP and two GTP molecules, this represents only a small fraction of the available energy released in the complete aerobic oxidation of glucose. The remaining energy is stored as high-energy electrons in the reduced coenzymes NADH and FADH2, which can be thought of as high-energy electron carriers. The goal of stages III and IV is to recover this energy in the form of ATP.