Transporters in the Inner Mitochondrial Membrane Help Maintain Appropriate Cytosolic and Matrix Concentrations of NAD+ and NADH
In the cytosol, NAD+ is required for step 6 of glycolysis (see Figure 12-3), and in the mitochondrial matrix, NAD+ is required for the conversion of pyruvate to acetyl CoA and for three steps in the citric acid cycle (step 4, step 5, and step 9 in Figure 12-16). In each case, NADH is a product of the reaction. If glycolysis and oxidation of pyruvate are to continue, NAD+ must be regenerated by oxidation of NADH to ensure that this substrate is available. (Similarly, the FADH2 generated in stage II reactions must be reoxidized to FAD if FAD-dependent reactions are to continue.) As we will see in the next section, the electron-transport chain within the inner mitochondrial membrane converts NADH to NAD+ and FADH2 to FAD as it reduces O2 to water and converts the energy stored in the high-energy electrons in the reduced forms of these molecules into a proton-motive force (stage III). Even though O2 is not involved in any reaction of the citric acid cycle, in the absence of O2 this cycle soon stops operating because in such anaerobic conditions, the mitochondria cannot regenerate the required NAD+ and FAD substrates. NAD+ and FAD dwindle due to the inability of the electron-transport chain within the mitochondrion to oxidize NADH and FADH2. These observations raise the question of how a supply of NAD+ in the cytosol is regenerated.
Page 536
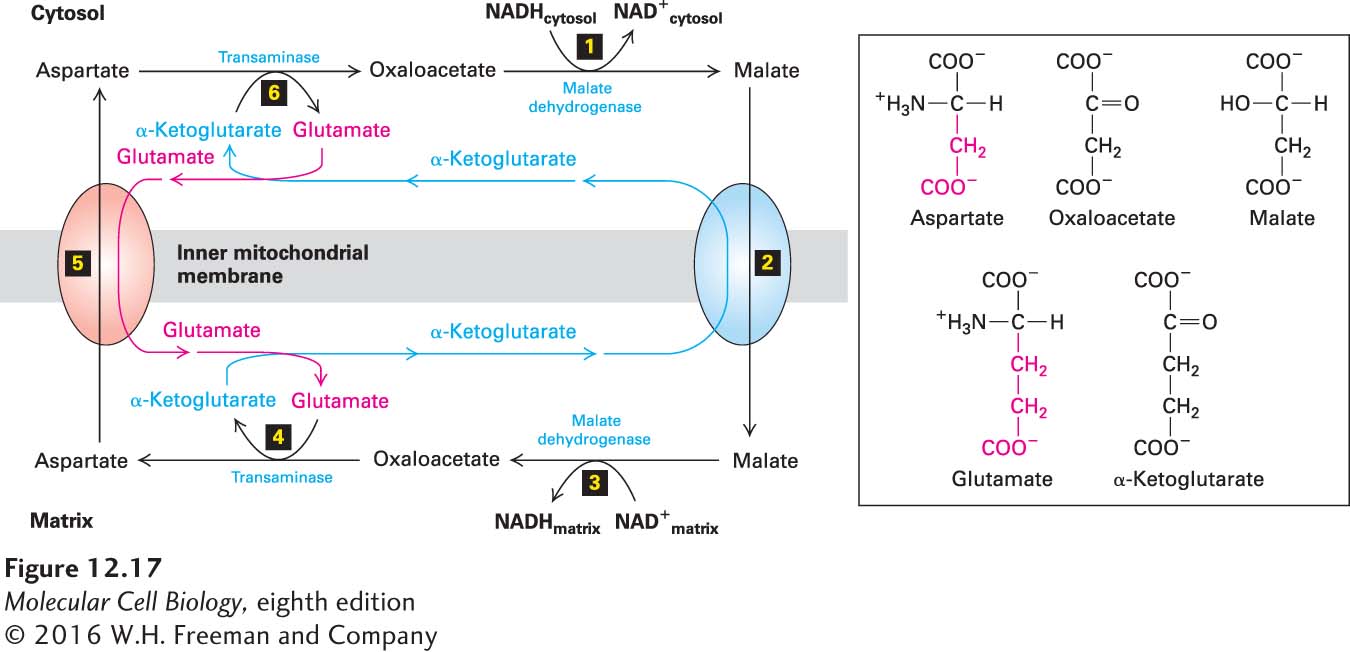
If the NADH from the cytosol could move into the mitochondrial matrix and be oxidized by the electron-transport chain, and if the NAD+ product could be transported back into the cytosol, regeneration of cytosolic NAD+ would be simple when O2 is available. However, the inner mitochondrial membrane is impermeable to NADH. To bypass this problem and permit the electrons from cytosolic NADH to be transferred indirectly to O2 via the mitochondrial electron-transport chain, cells use several electron shuttles to transfer electrons from NADH in the cytoplasm to NAD+ in the matrix. The operation of the most widespread shuttle—the malate-aspartate shuttle—is depicted in Figure 12-17.
FIGURE 12-17The malate-aspartate shuttle. This cyclical series of reactions transfers electrons from NADH in the cytosol (via the intermembrane space) across the inner mitochondrial membrane, which is impermeable to NADH itself, to NAD+ in the matrix. The net result is the replacement of cytosolic NADH with NAD+ and matrix NAD+ with NADH. Step 1: Cytosolic malate dehydrogenase transfers electrons from cytosolic NADH to oxaloacetate, forming malate. Step 2: An antiporter (blue oval) in the inner mitochondrial membrane transports malate into the matrix in exchange for α-ketoglutarate. Step 3: Mitochondrial malate dehydrogenase converts malate back to oxaloacetate, reducing NAD+ in the matrix to NADH in the process. Step 4: Oxaloacetate, which cannot directly cross the inner membrane, is converted to aspartate by addition of an amino group from glutamate. In this transaminase-catalyzed reaction in the matrix, glutamate is converted to α-ketoglutarate. Step 5: A second antiporter (red oval) exports aspartate to the cytosol in exchange for glutamate. Step 6 A cytosolic transaminase converts aspartate to oxaloacetate and α-ketoglutarate to glutamate, completing the cycle. The blue arrows reflect the movement of the α-ketoglutarate, the red arrows the movement of glutamate, and the black arrows that of aspartate/malate. It is noteworthy that as aspartate and malate cycle clockwise, glutamate and α-ketoglutarate cycle in the opposite direction.
For every complete cycle of the shuttle, there is no overall change in the numbers of NADH and NAD+ molecules or the intermediates aspartate or malate. In the cytosol, however, NADH is oxidized to NAD+, which can be used for glycolysis, and in the matrix, NAD+ is reduced to NADH, which can be used for electron transport: