Electron Transport in Mitochondria Is Coupled to Proton Pumping
During electron transport from NADH and FADH2 to O2, protons from the mitochondrial matrix are pumped across the inner membrane. This pumping raises the pH of the mitochondrial matrix relative to the intermembrane space and cytosol and also makes the matrix more electrically negative with respect to the intermembrane space. In other words, the free energy released during the oxidation of NADH or FADH2 is stored both as a proton concentration gradient and as an electrical gradient across the membrane—collectively known as the proton-motive force (see Figure 12-2). As we will see in Section 12.5, the movement of protons back across the inner membrane, driven by this force, is coupled to the synthesis of ATP from ADP and Pi by ATP synthase (stage IV).
The synthesis of ATP from ADP and Pi, driven by the energy released by transfer of electrons from NADH or FADH2 to O2, is the major source of ATP in aerobic non-photosynthetic cells. Much evidence shows that in mitochondria and bacteria, this process of oxidative phosphorylation depends on the generation of a proton-motive force across the inner membrane (in mitochondria) or bacterial plasma membrane, with electron transport, proton pumping, and ATP formation occurring simultaneously. In the laboratory, for instance, addition of O2 and an oxidizable substrate such as pyruvate or succinate to isolated intact mitochondria results in net synthesis of ATP if the inner mitochondrial membrane is intact. In the presence of minute amounts of detergents that make the membrane leaky, electron transport and the oxidation of these metabolites by O2 still occurs. However, no ATP is made under these conditions because the proton leak prevents the maintenance of the proton-motive force.
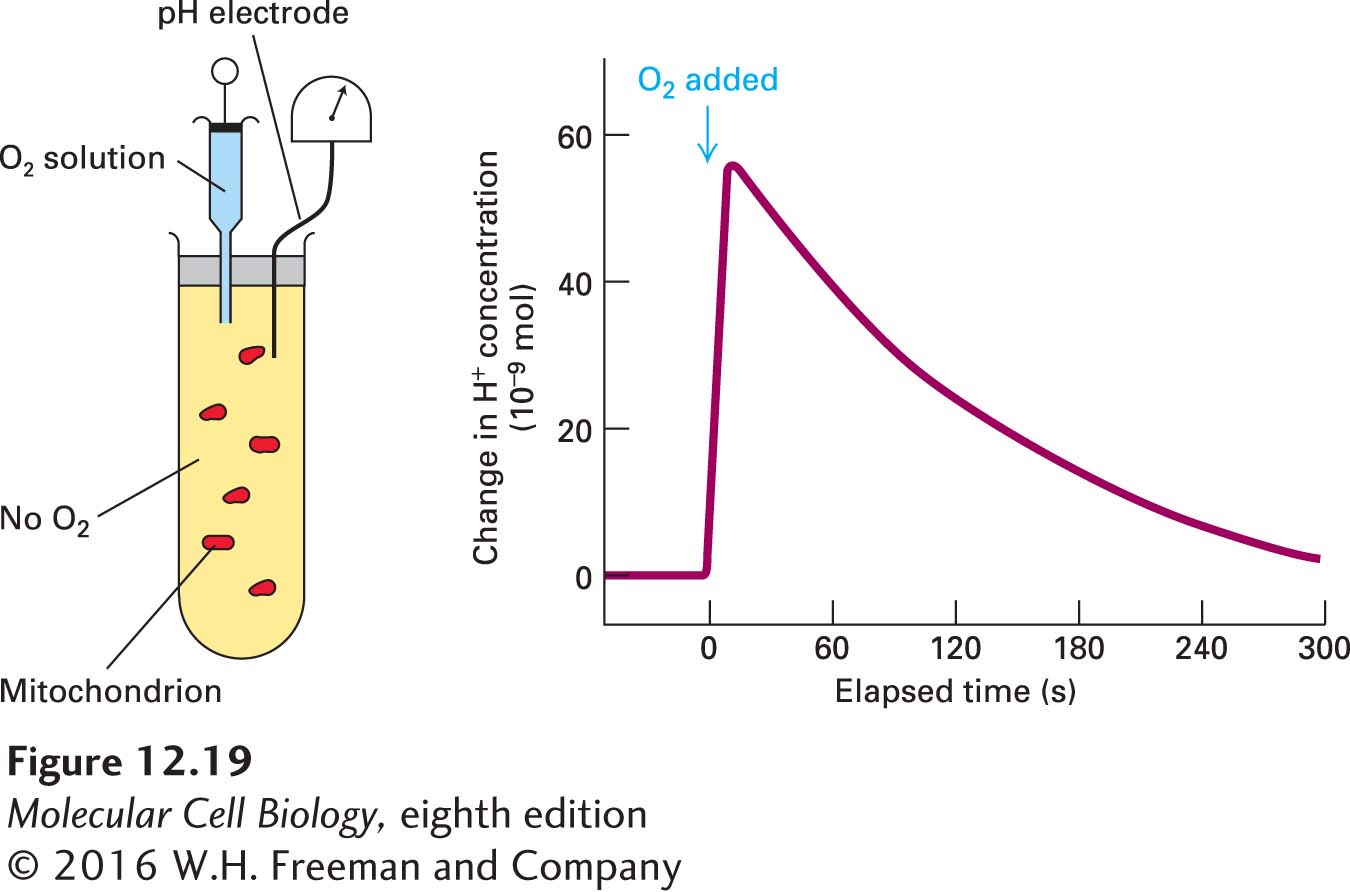
The coupling between electron transport from NADH (or FADH2) to O2 and proton transport across the inner mitochondrial membrane can be demonstrated experimentally with isolated, intact mitochondria (Figure 12-19). As soon as O2 is added to a suspension of mitochondria in an otherwise O2-free solution that contains NADH, the medium outside the mitochondria transiently becomes more acidic (increased proton concentration) because the mitochondrial outer membrane is freely permeable to protons. (Remember that the malate-aspartate shuttle and other shuttles can convert the NADH in the solution into NADH in the mitochondrial matrix.) Once the O2 is depleted by its reduction, the excess protons in the medium slowly leak back into the matrix. By measuring the pH change in such experiments, one can calculate that about 10 protons are transported out of the matrix for every electron pair transferred from NADH to O2.
EXPERIMENTAL FIGURE 12-19Electron transfer from NADH to O2 is coupled to proton transport across the mitochondrial membrane. If NADH is added to a suspension of mitochondria depleted of O2, no NADH is oxidized. When a small amount of O2 is added to the system (arrow), there is a sharp rise in the concentration of protons in the surrounding medium outside the mitochondria (decrease in pH). Thus the oxidation of NADH by O2 is coupled to the movement of protons out of the matrix. Once the O2 is depleted, the excess protons slowly move back into the mitochondria (powering the synthesis of ATP), and the pH of the extracellular medium returns to its initial value.
Page 540
To obtain numbers for FADH2, the above experiment can be repeated using succinate instead of NADH as the substrate. (Recall that oxidation of succinate to fumarate in the citric acid cycle generates FADH2; see Figure 12-16.) The amount of succinate added can be adjusted so that the amount of FADH2 generated is equivalent to the amount of NADH in the first experiment. As in the first experiment, addition of O2 causes the medium outside the mitochondria to become acidic, but less so than with NADH. This difference is not surprising, because electrons in FADH2 have less potential energy (43.4 kcal/mol) than electrons in NADH (52.6 kcal/mol), and thus FADH2 drives the translocation of fewer protons from the matrix and a smaller change in pH.