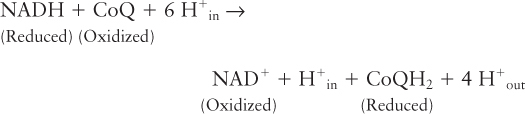Four Large Multiprotein Complexes Couple Electron Transport to Proton Pumping Across the Inner Mitochondrial Membrane
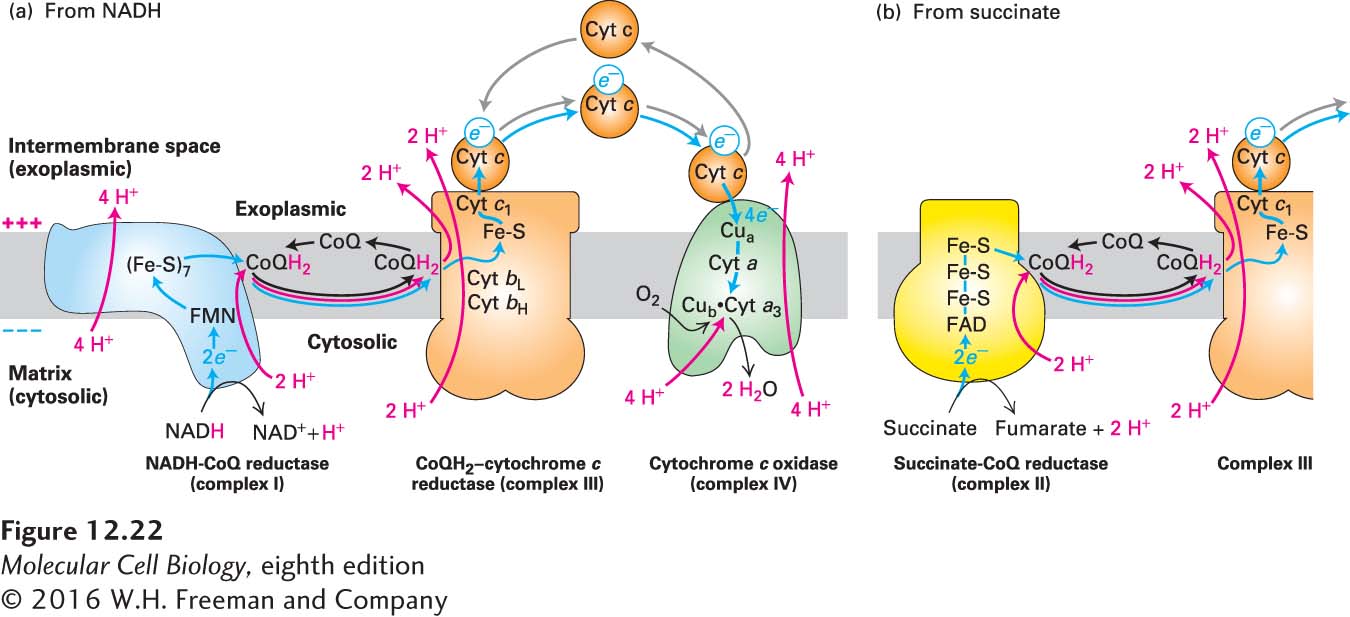
As electrons flow downhill from one electron carrier to the next in the electron-transport chain, the energy released is used to power the pumping of protons against their electrochemical gradient across the inner mitochondrial membrane. Four large multiprotein complexes (Figure 12-22) directly or indirectly couple the movement of electrons to proton pumping: NADH-CoQ reductase (complex I, >40 subunits), succinate-CoQ reductase (complex II, 4 subunits), CoQH2–cytochrome c reductase (complex III, 11 subunits), and cytochrome c oxidase (complex IV, 13 subunits). The electrons follow one of two routes through these complexes: I → III → IV or II → III → IV. Complexes I, III, and IV all pump protons directly across the inner membrane, whereas complex II does not (see Figure 12-22). Electrons from NADH flow from complex I via CoQ/CoQH2 to complex III and then, via the soluble protein cytochrome c (cyt c), to complex IV to reduce molecular oxygen (complex II is bypassed) (see Figure 12-22a); electrons from FADH2 flow from complex II via CoQ/CoQH2 to complex III (see Figure 12-22b) and then via cytochrome c to complex IV to reduce molecular oxygen (complex I is bypassed).
FIGURE 12-22The mitochondrial electron-transport chain. Electrons (blue arrows) flow through four major multiprotein complexes (I–IV). Electron movement between complexes is mediated either by the lipid-soluble molecule coenzyme Q (CoQ, oxidized form; CoQH2, reduced form) or the water-soluble protein cytochrome c (cyt c). Three of the multiprotein complexes use the energy released from the electrons to pump protons (red arrows) from the matrix (cytosolic compartment in bacteria) to the intermembrane space (exoplasmic space of bacteria). (a) Pathway from NADH. Electrons from NADH (2/NADH oxidized) flow through complex I, initially via a flavin mononucleotide (FMN) and then sequentially via seven iron-sulfur clusters (Fe-S), to CoQ, to which two protons bind, forming CoQH2. Conformational changes in complex I that accompany the electron flow drive proton pumping from the matrix to the intramembrane space. Electrons then flow via the released (and subsequently recycled) CoQH2 to complex III, and then via cyt c to complex IV. Each of four cyt c molecules carrying four electrons originating from two NADHs transfers individual electrons to complex IV for the four-electron reduction of one O2 molecule to two H2O molecules and transport of four protons. Thus, for every two NADHs oxidized and one O2 molecule reduced, a total of 20 protons are translocated out of the matrix into the intermembrane space. (b) Pathway from succinate. Two electrons flow from each succinate to complex II via FAD/FADH2 and iron-sulfur clusters (Fe-S), from complex II to complex III via CoQ/CoQH2, and then to complex IV via cyt c. Electrons released during oxidation of succinate to fumarate in complex II are used to reduce CoQ to CoQH2 without translocating additional protons. The remainder of electron transport from CoQH2 proceeds by the same pathway as for the NADH pathway in (a). Thus, for every two succinates oxidized and one O2 molecule reduced,12 protons (8 via the Q-cycle of complex III, 4 via complex IV) are translocated.
As shown in Figure 12-22, CoQ accepts electrons released from NADH-CoQ reductase (complex I) or succinate-CoQ reductase (complex II) and donates them to CoQH2–cytochrome c reductase (complex III). Protons are simultaneously transported from the matrix side of the membrane (also called the cytosolic side) to the intermembrane space (also called the exoplasmic side). Whenever CoQ accepts electrons, it does so at a binding site on the matrix side of a protein complex, always picking up protons from the medium there. Whenever CoQH2 releases its electrons, it does so at a site on the intermembrane space side of a protein complex, releasing protons into the fluid of the intermembrane space. Thus the transport of each pair of electrons by CoQ is obligately coupled to the movement of two protons from the matrix to the intermembrane space.
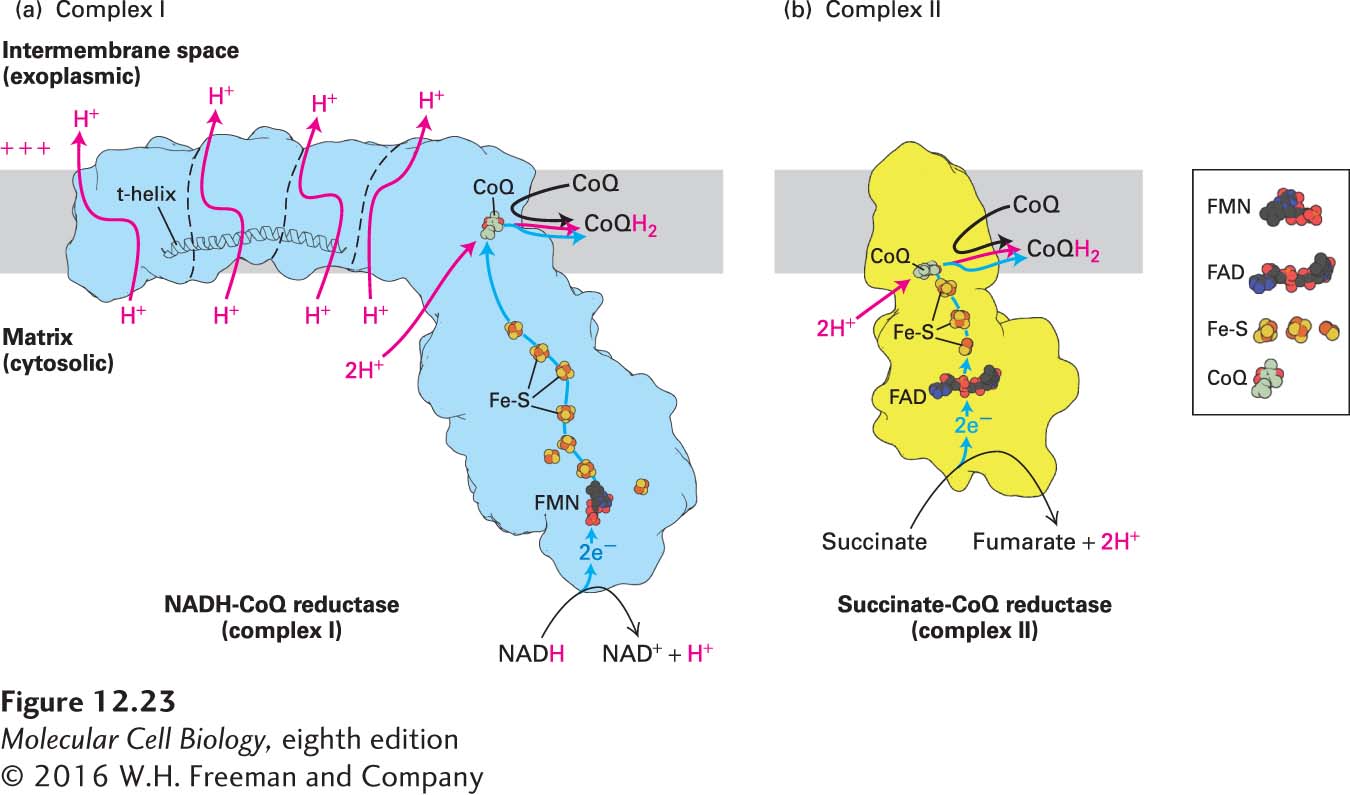
NADH-CoQ Reductase (Complex I) Electrons are transferred from NADH to CoQ by NADH-CoQ reductase (see Figure 12-22a). Electron microscopy and x-ray crystallography of complex I from bacteria (mass ~500 kDa, with 14 subunits) and from eukaryotes (~1 MDa, with 14 highly conserved core subunits shared with bacteria plus about 26–32 accessory subunits) has established that it is L-shaped (Figure 12-23a). The membrane-embedded arm of the L is slightly curved, about 180 Å long, and comprises proteins with more than 60 transmembrane α helices. This arm has four subdomains, three of which contain proteins that are members of a family of cation antiporters. The hydrophilic peripheral arm extends over 130 Å away from the membrane into the matrix (cytosolic) space.
Page 543
[Part (a) data from V. Zickermann et al., 2015, Science347:44–49, PDB ID 3m9s. Part (b) data from F. Sun et al., 2005, Cell121:1043–1057, PDB ID 1zoy.]
FIGURE 12-23Electron and proton transport through complexes I and II. (a) Model of complex I based on its three-dimensional structure. The outline of the shape of the core complex I from the yeast Y. lipolytica, as determined by x-ray crystallography, is shown in light blue, and the borders separating several distinct structural subunits are indicated by thin dashed black lines. From NADH, electrons flow first to a flavin mononucleotide (FMN) and then, via iron-sulfur clusters (Fe-S, red and yellow balls), to CoQ, to which two protons from the matrix bind (red arrow) to form CoQH2. Conformational changes due to the electron flow, which may be induced by changes in the charge of the CoQ and may include a piston-like horizontal movement of the t-helix, drive proton pumping through the transmembrane subunits from the matrix to the intramembrane space (red arrows). (b) Model of complex II based on its three-dimensional structure. Electrons flow through complex II from succinate to CoQ via FAD/FADH2 and iron-sulfur clusters (Fe-S), and from complex II to complex III via CoQ/CoQH2. Electrons released during oxidation of succinate to fumarate in complex II are used to reduce CoQ to CoQH2 without translocating additional protons.
[Part (a) data from V. Zickermann et al., 2015, Science347:44–49, PDB ID 3m9s. Part (b) data from F. Sun et al., 2005, Cell121:1043–1057, PDB ID 1zoy.]
NAD+ is exclusively a two-electron carrier: it accepts or releases a pair of electrons simultaneously. In NADH-CoQ reductase, the NADH-binding site is at the tip of the peripheral arm (see Figure 12-23a); electrons released from NADH first flow to FMN (flavin mononucleotide), a prosthetic group, or cofactor, related to FAD, then are shuttled about 95 Å down the peripheral arm through a series of iron-sulfur clusters and finally to CoQ, which is bound at a site at least partially in the plane of the membrane. FMN, like FAD, can accept two electrons, but does so one electron at a time.
Each transported electron undergoes a drop in potential of about 360 mV, equivalent to a ΔG°′ of −16.6 kcal/mol for the two electrons transported. Much of this released energy is used to transport four protons across the inner membrane per molecule of NADH oxidized by complex I. Those four protons are distinct from the two protons that are transferred to the CoQ as illustrated in Figures 12-21, 12-22a, and 12-23a. The precise mechanism by which the energy released by electron transport in the peripheral arm is used to change the conformation of subunits in the membrane arm and thus mediate the movement of four protons across the membrane is uncertain. Three protons are likely to pass through the three cation antiporter domains via a zigzag series of polar side chains that can be protonated and that span the membrane. A similar series of side chains is the likely conduit for the fourth proton. A transverse α helix (t-helix) in the membrane arm runs parallel to the plane of the membrane, potentially mechanically linking the antiporter-like domains to the peripheral arm (see Figure 12-23a) and possibly contributing directly to the conformational changes required to convert the energy released by the electron transport in the peripheral arm into proton transport by the transmembrane domains.
The overall reaction catalyzed by this complex is
Page 544
Succinate-CoQ Reductase (Complex II) Succinate dehydrogenase, the enzyme that oxidizes a molecule of succinate to fumarate in the citric acid cycle (and in the process generates the reduced coenzyme FADH2), is one of the four subunits of complex II (Figure 12-23b). Thus the citric acid cycle is physically as well as functionally linked to the electron-transport chain. The two electrons released in the conversion of succinate to fumarate are transferred first to FAD in succinate dehydrogenase, then to iron-sulfur clusters—regenerating FAD—and finally to CoQ, which binds to a cleft on the matrix side of the transmembrane portions of complex II (see Figures 12-22b and 12-23b). The pathway is somewhat reminiscent of that in complex I (see Figure 12-23a).
The overall reaction catalyzed by this complex is
Although the ΔG°′ for this reaction is negative, the released energy is insufficient for proton pumping in addition to reduction of CoQ to form CoQH2. Thus no protons are translocated directly across the membrane by succinate-CoQ reductase, and no proton-motive force is generated in this part of the electron-transport chain. We will see shortly how the protons and electrons in the CoQH2 molecules generated by complexes I and II contribute to the generation of the proton-motive force.
Complex II generates CoQH2 from succinate via FAD/FADH2-mediated redox reactions. Another set of proteins in the matrix and inner mitochondrial membrane performs a comparable set of FAD/FADH2-mediated redox reactions, producing CoQH2 from fatty acid oxidation and contributing electrons to the electron-transport chain (see Figure 12-18). Fatty acyl–CoA dehydrogenase, which is a water-soluble enzyme, catalyzes the first step of the oxidation of fatty acyl CoA in the mitochondrial matrix. There are several fatty acyl–CoA dehydrogenase enzymes with specificities for fatty acyl chains of different lengths. These enzymes mediate the initial step in a four-step process that removes two carbons from the fatty acyl group by oxidizing the carbon in the β position of the fatty acyl chain (thus the entire process is often referred to as β-oxidation). These reactions generate acetyl CoA, which in turn enters the citric acid cycle. They also generate an FADH2 intermediate and NADH. The FADH2 generated remains bound to the enzyme during the redox reaction, as is the case for complex II. A water-soluble protein called electron transfer flavoprotein (ETF) transfers the high-energy electrons from the FADH2 in the fatty acyl–CoA dehydrogenase to electron transfer flavoprotein:ubiquinone oxidoreductase (ETF:QO), a membrane protein that reduces CoQ to CoQH2 in the inner membrane. This CoQH2 intermixes in the membrane with the other CoQH2 molecules generated by complexes I and II, all contributing to proton transport out of the matrix by complex III.
CoQH2–Cytochrome c Reductase (Complex III) A CoQH2 generated by complex I, complex II, or ETF:QO donates two electrons to CoQH2–cytochrome c reductase (complex III), regenerating oxidized CoQ. Concomitantly, it releases into the intermembrane space two protons previously picked up by CoQ on the matrix face, generating part of the proton-motive force (see Figure 12-22). Within complex III, the released electrons are first transferred to an iron-sulfur cluster within the complex and then to cytochrome c1 or to two b-type cytochromes (bL and bH, see the description of the Q cycle below). Finally, the two electrons are transferred sequentially to two molecules of the oxidized form of cytochrome c, a water-soluble peripheral protein that diffuses in the intermembrane space. For each pair of electrons transferred, the overall reaction catalyzed by complex III is
The ΔG°′ for this reaction is sufficiently negative that two protons in addition to those from CoQH2 are translocated from the mitochondrial matrix across the inner membrane for each pair of electrons transferred; this transfer involves the proton-motive Q cycle, discussed below. The heme protein cytochrome c and the small lipid-soluble molecule CoQ play similar roles in the electron-transport chain in that they both serve as mobile electron shuttles, transferring electrons (and thus energy) between the complexes of the electron-transport chain.
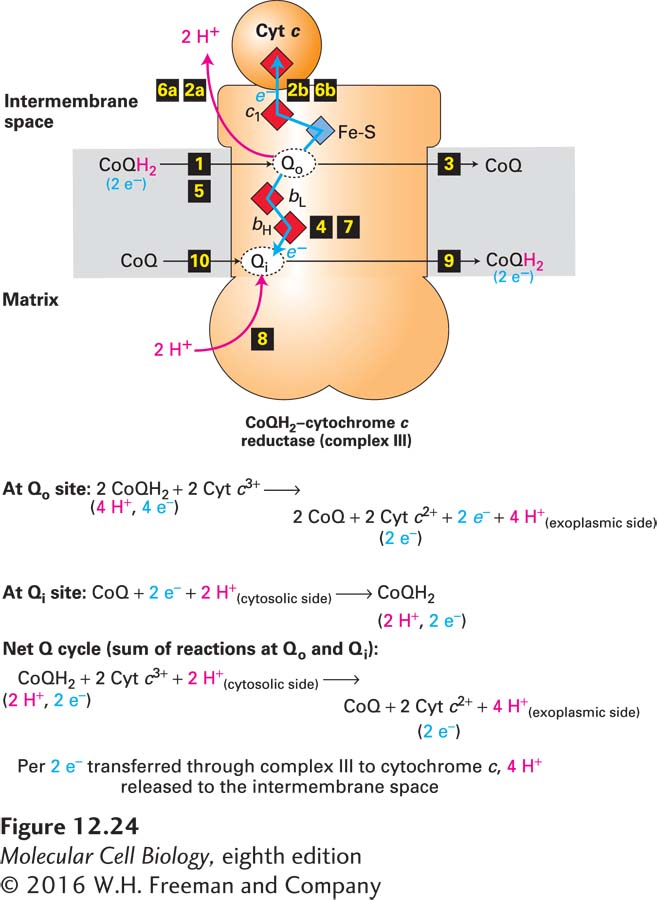
The Q Cycle Experiments have shown that four protons are translocated across the inner mitochondrial membrane per electron pair transported from CoQH2 through complex III. These four protons are those carried on two CoQH2 molecules, which are converted to two CoQ molecules during the cycle. However, another CoQ molecule receives two other protons from the matrix and is converted to one CoQH2 molecule. Thus the net overall reaction involves the conversion of only one CoQH2 molecule to CoQ as two electrons are transferred, one at a time, to two molecules of the acceptor cytochrome c. An evolutionarily conserved mechanism, called the Q cycle, is responsible for the two-for-one transport of protons and electrons by complex III (Figure 12-24).
FIGURE 12-24The Q cycle. The Q cycle of complex III uses the net oxidation of one CoQH2 molecule to transfer four protons into the intermembrane space and two electrons to two cytochrome c molecules. The cycle begins when a molecule from the combined pool of reduced CoQH2 in the inner mitochondrial membrane binds to the Qo site on the intermembrane space (exoplasmic) side of the transmembrane portion of complex III (step 1). There CoQH2 releases two protons into the intermembrane space (step 2a), and two electrons and the resulting CoQ dissociate (step 3). One of the electrons is transported, via an iron-sulfur protein and cytochrome c1, directly to cytochrome c (step 2b). (Recall that each cytochrome c shuttles one electron from complex III to complex IV.) The other electron moves through cytochromes bL and bH and partially reduces an oxidized CoQ molecule bound to the second, Qi, site on the matrix (cytosolic) side of the complex, forming a CoQ semiquinone anion, Q•− (step 4). The process is repeated with the binding of a second CoQH2 at the Qo site (step 5), proton release (step 6a), reduction of another cytochrome c (step 6b), and addition of the other electron to the Q•− bound at the Qi site (step 7). There the addition of two protons from the matrix yields a fully reduced CoQH2 molecule at the Qi site, which then dissociates (steps 8 and 9), freeing the Qi to bind a new molecule of CoQ (step 10) and begin the Q cycle over again. See B. Trumpower, 1990, J. Biol. Chem.265:11409, and E. Darrouzet et al., 2001, Trends Biochem. Sci. 26:445.
The substrate for complex III, CoQH2, is generated by several enzymes, including NADH-CoQ reductase (complex I), succinate-CoQ reductase (complex II), ETF:QO (during β-oxidation), and as we shall see, by complex III itself.
As shown in Figure 12-24, in one turn of the Q cycle, two molecules of CoQH2 are oxidized to CoQ at the Qo site in complex III and release a total of four protons into the intermembrane space, but at the Qi site, one molecule of CoQH2 is regenerated from CoQ and two additional protons from the matrix. The translocated protons are all derived from CoQH2, which obtained its protons from the matrix, as described above. Although seemingly cumbersome, the Q cycle optimizes the number of protons pumped per pair of electrons moving through complex III. The Q cycle is found in all plants and animals as well as in bacteria. Its formation at a very early stage of cellular evolution was probably essential for the success of all life forms as a way of converting the potential energy in reduced coenzyme Q into the maximum proton-motive force across a membrane. In turn, this process maximizes the number of ATP molecules synthesized from each electron that moves down the electron-transport chain from NADH or FADH2 to O2.
Page 545
How are the two electrons released from CoQH2 at the Qo site directed to different acceptors, either to Fe-S, cytochrome c1, and then cytochrome c (upward pathway in Figure 12-24), or alternatively to cytochrome bL, cytochrome bH, and then CoQ at the Qi site (downward pathway in Figure 12-24)? The mechanism involves a flexible hinge in the Fe-S–containing protein subunit of complex III. Initially, the Fe-S cluster is close enough to the Qo site to pick up an electron from CoQH2 bound there. Once this happens, the subunit containing this Fe-S cluster swings the cluster away from the Qo site to a position near enough to the heme on cytochrome c1 for electron transfer to occur. With the Fe-S–containing subunit in this alternate conformation, the second electron released from the CoQH2 bound to the Qo site cannot move to the Fe-S cluster—it is too far away, so it takes an alternative path open to it via a somewhat less thermodynamically favored route to cytochrome bL and through cytochrome bH to the CoQ at the Qi site.
Cytochrome c Oxidase (Complex IV) Cytochrome c, after being reduced by one electron from complex III, is reoxidized as it transports its electron to cytochrome c oxidase (complex IV) (see Figure 12-22a). Mitochondrial cytochrome c oxidases contain 13 different subunits, but the catalytic core of the enzyme consists of only three. The functions of the remaining subunits are not well understood. Bacterial cytochrome c oxidases contain only the three catalytic subunits. In both mitochondria and bacteria, four molecules of reduced cytochrome c bind, one at a time, to the oxidase. An electron is transferred from the heme of each cytochrome c, first to the pair of copper ions labeled Cua2+, then to the heme in cytochrome a, and next to the oxygen reduction center, composed of Cub2+ and the heme in cytochrome a3, which together bind in a sandwich fashion and sequester the O2 molecule that will be reduced to two H2Os. Several lines of evidence suggest that O2 reaches the reduction center via one or more hydrophobic channels through the protein, into which O2 flows from the hydrophobic core of the membrane, where it is more soluble than in aqueous solution (see Figure 12-22a).
The four electrons are finally passed by complex IV to O2, the ultimate electron acceptor, yielding two H2O, which together with CO2 is one of the end products of the overall oxidation pathway. Proposed intermediates in oxygen reduction include the peroxide anion (O22−) and the hydroxyl radical (OH•), as well as unusual complexes of iron and oxygen atoms. These intermediates would be harmful to the cell if they escaped from complex IV, but they do so only rarely (see the discussion of reactive oxygen species below) because they are sequestered by the Cub2+ and the heme in cytochrome a3. To generate H2O from the reduced O2, protons are channeled to the reduction center. In addition, other protons are transported across the membrane from the matrix to the intermembrane space. During the transport of four electrons through the cytochrome c oxidase complex, four protons from the matrix are translocated across the membrane. Thus complex IV transports only one proton per electron transferred, whereas complex III, using the Q cycle, transports two protons per electron transferred. However, the mechanism by which complex IV translocates these protons and energetically couples the translocation to O2 reduction is not fully understood.
Page 546
For each four electrons transferred, the overall reaction catalyzed by cytochrome c oxidase is
The poison cyanide, which has been used as a chemical warfare agent, by spies to commit suicide when captured, in gas chambers to execute prisoners, and by the Nazis (Zyklon B gas) for the mass murder of Jews and others, is toxic because it binds to the heme a3 in mitochondrial cytochrome c oxidase (complex IV), inhibiting electron transport and thus oxidative phosphorylation and production of ATP. Cyanide is one of many toxic small molecules that interfere with energy production in mitochondria.