ATP Synthase Comprises F0 and F1 Multiprotein Complexes
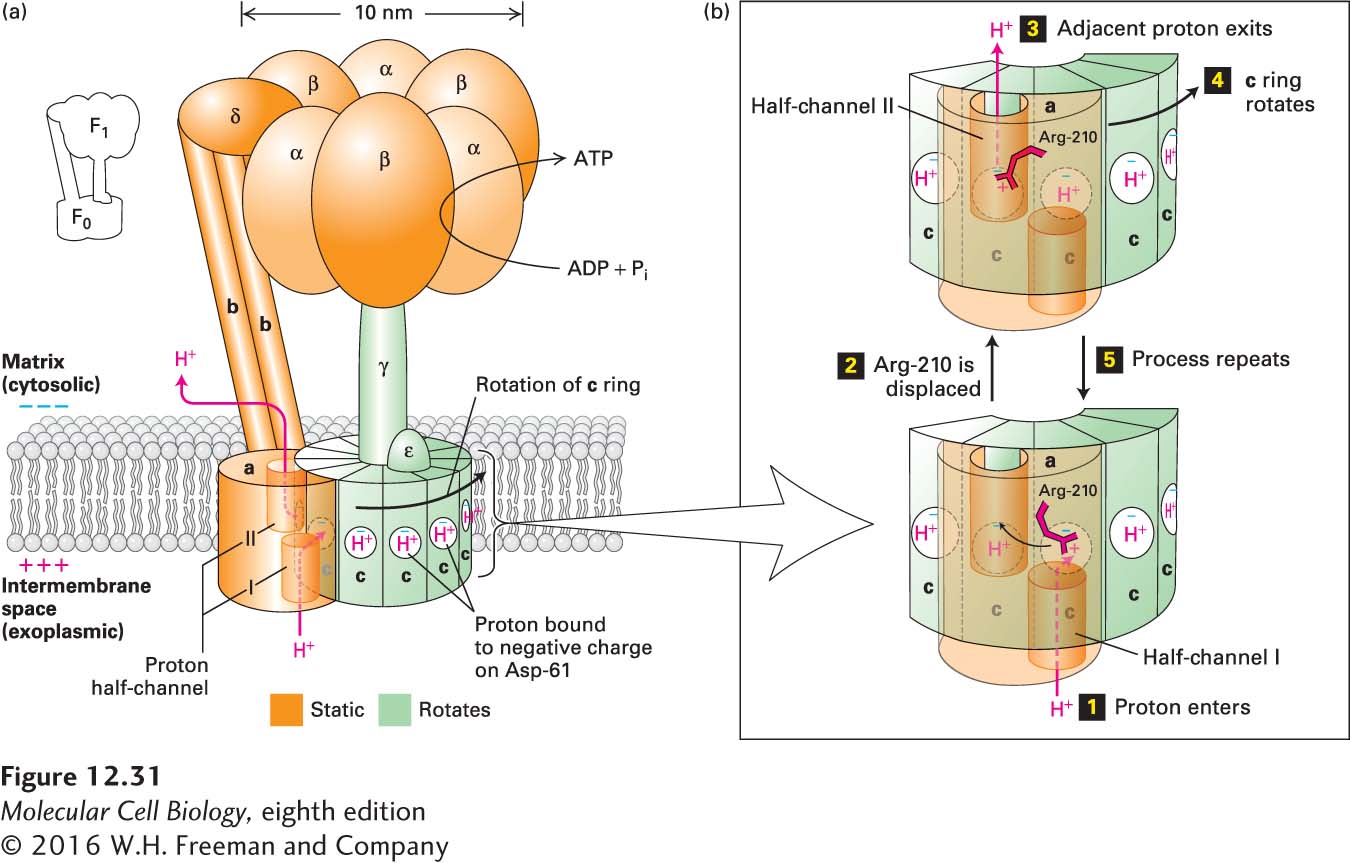
With general acceptance of Mitchell’s chemiosmotic mechanism, researchers turned their attention to the structure and operation of ATP synthase. The complex has two principal subcomplexes, F0 and F1, both of which are multimeric proteins (Figure 12-31a). The F0 subcomplex contains three types of integral membrane proteins, designated a, b, and c. In bacteria and in yeast mitochondria, the most common subunit stoichiometry is a1b2c10, but the number of c subunits per subcomplex varies among different eukaryotes from 8 to 15. In all cases, the c subunits form a doughnut-shaped ring (“c ring”) in the plane of the membrane. The one a and two b subunits are rigidly linked to one another, but not to the c ring, a critical feature of the protein to which we will return shortly.
FIGURE 12-31Structure of ATP synthase (the F0F1 complex) in the bacterial plasma membrane and mechanism of proton translocation across the membrane. (a) The F0 membrane-embedded subcomplex of ATP synthase is built of three integral membrane proteins: one copy of a, two copies of b, and an average of ten copies of c arranged in a ring in the plane of the membrane. Two proton half-channels near the interfaces of subunit a with the c subunits mediate proton movement across the membrane (proton path is indicated by red arrows). Half-channel I allows protons to move one at a time from the exoplasmic medium (equivalent to intermembrane space in mitochondria) to the negatively charged side chain of Asp-61 in the center of a c subunit near the middle of the membrane. The proton-binding site in each c subunit is represented as a white circle with a blue “−” representing the negative charge on the side chain of Asp-61. Half-channel II permits protons to move from the Asp-61 of an adjacent c subunit into the cytosolic medium. The detailed structure of the c ring and a portion of the adjacent a subunit is shown in Figure 12-34. The F1 subcomplex of ATP synthase contains three copies each of subunits α and β, which form a hexamer resting atop the single rod-shaped γ subunit, which is inserted into the c ring of F0. The ε subunit is rigidly attached to the γ subunit and also to several of the c subunits. The δ subunit permanently links one of the α subunits in the F1 subcomplex to the b subunit of F0. Thus the F0a and b subunits and the F1 δ subunit and (αβ)3 hexamer form a rigid structure (orange) anchored in the membrane. During proton flow, the c ring and the attached F1 ε and γ subunits rotate as a unit (green), causing conformational changes in the F1 β subunits, leading to ATP synthesis. (b) Potential mechanism of proton translocation. Step 1: A proton from the exoplasmic space enters half-channel I and moves toward the “empty” (unprotonated) Asp-61 proton-binding site. The negative charge (blue “−”) on the unprotonated side chain Asp-61 is balanced, in part, by a positive charge on the side chain of Arg-210 (red “+”). Step 2: The proton fills the empty proton-binding site and simultaneously displaces the positively charged Arg-210 side chain, which swings over to the filled proton-binding site on the adjacent c subunit (curved arrow). As a consequence, the proton bound at that adjacent site is displaced. Step 3: The displaced adjacent proton moves through half-channel II and is released into the cytosolic space, leaving an empty proton-binding site on Asp-61. Step 4: Counterclockwise rotation of the entire c ring moves the “empty” c subunit over half-channel I. Step 5: The process is repeated. See M. J. Schnitzer, 2001, Nature410:878; P. D. Boyer, 1999, Nature402:247; and C. von Ballmoos, A. Wiedenmann, and P. Dimroth, 2009, Annu. Rev. Biochem.78:649.
The F1 subcomplex is a water-soluble complex of five distinct polypeptides with the composition α3β3γδε that is normally firmly bound to the F0 subcomplex at the surface of the membrane. The lower end of the rodlike γ subunit of the F1 subcomplex is a coiled coil that fits into the center of the c-subunit ring of F0 and appears rigidly attached to it. Thus when the c-subunit ring rotates, the rodlike γ subunit moves with it. The F1 ε subunit is rigidly attached to γ and also forms tight contacts with several of the c subunits of F0. The α and β subunits are responsible for the overall globular shape of the F1 subcomplex and associate in alternating order to form a hexamer, αβαβαβ, or (αβ)3, which rests atop the single long γ subunit. The F1 δ subunit is permanently linked to one of the F1 α subunits and also binds to the b subunit of F0. Thus the a and b subunits of the F0 subcomplex and the δ subunit and (αβ)3 hexamer of the F1 subcomplex form a rigid structure anchored in the membrane. The rodlike b subunits form a “stator” that prevents the (αβ)3 hexamer from moving while it rests on the γ subunit, whose rotation, together with that of the c subunits of F0, plays an essential role in the ATP synthesis mechanism described below.
Page 554
When ATP synthase is embedded in a membrane, the F1 subcomplex forms a knob that protrudes from the cytosolic face (the matrix face in the mitochondrion). Because F1 separated from membranes is capable of catalyzing ATP hydrolysis (ATP conversion to ADP plus Pi) in the absence of the F0 subcomplex, it has been called the F1 ATPase; however, its function in cells is the reverse, to synthesize ATP. ATP hydrolysis is a spontaneous process (ΔG < 0); thus energy is required to drive the ATPase “in reverse” and generate ATP.