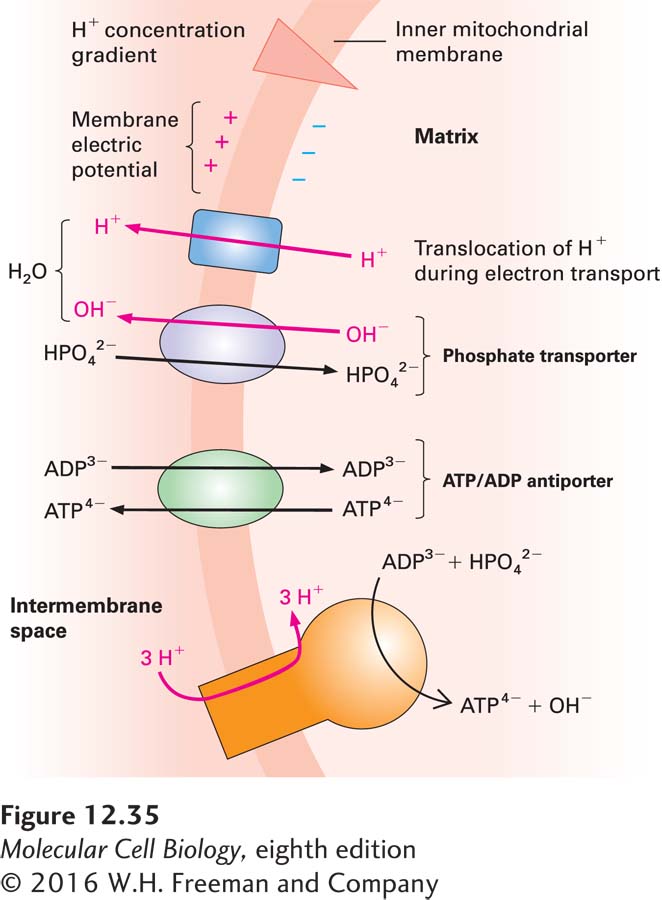
ATP-ADP Exchange Across the Inner Mitochondrial Membrane Is Powered by the Proton-Motive Force
The proton-motive force is used to power multiple energy-requiring processes in cells. In addition to powering ATP synthesis, the proton-motive force across the inner mitochondrial membrane powers the exchange of ATP formed by oxidative phosphorylation inside the mitochondrion for ADP and Pi in the cytosol. This exchange, which is required to supply ADP and Pi substrates for oxidative phosphorylation to continue, is mediated by two proteins in the inner membrane: a phosphate transporter (HPO42−/OH− antiporter), which mediates the import of one HPO42− coupled to the export of one OH−, and an ATP/ADP antiporter (Figure 12-35). The ATP/ADP antiporter allows one molecule of ADP to enter the matrix only if one molecule of ATP exits simultaneously. The ATP/ADP antiporter, a dimer of two 30,000-Da subunits, makes up 10–15 percent of the protein in the inner mitochondrial membrane, so it is one of the more abundant mitochondrial proteins. The functioning of the two antiporters together produces an influx of one ADP3− and one Pi2− and an efflux of one ATP4− together with one OH−. Each OH− transported outward combines with a proton, translocated during electron transport to the intermembrane space, to form H2O. Thus proton translocation via electron transport drives the overall reaction in the direction of ATP export and ADP and Pi import.
FIGURE 12-35The phosphate and ATP/ADP transport system in the inner mitochondrial membrane. The coordinated action of two antiporters (purple and green), which results in the uptake of one ADP3− and one HPO42− in exchange for one ATP4− and one hydroxyl, is powered by the outward translocation of one proton (mediated by the proteins of the electron-transport chain, blue) during electron transport. The outer membrane is not shown here because it is permeable to molecules smaller than 5000 Da.
Page 557
Page 558
Because some of the protons translocated out of the mitochondrion during electron transport provide the power (by combining with the exported OH−) for the ATP-ADP exchange, fewer protons are available for ATP synthesis. It is estimated that for every four protons translocated out, three are used to synthesize one ATP molecule and one is used to power the export of ATP from the mitochondrion in exchange for ADP and Pi. This expenditure of energy from the proton concentration gradient to export ATP from the mitochondrion in exchange for ADP and Pi ensures a high ratio of ATP to ADP in the cytosol, where hydrolysis of the high-energy phosphoanhydride bond of ATP is used to power many energy-requiring reactions.
Studies of what turned out to be ATP/ADP antiporter activity were first recorded about 2000 years ago, when Dioscorides (~AD 40–90) described the effects of a poisonous herb from the thistle Atractylis gummifera, found commonly in the Mediterranean region. The same agent is found in the traditional Zulu multipurpose herbal remedy impila (Callilepis laureola). In Zulu, impila means “health,” although this herb has been associated with numerous poisonings. In 1962, the active agent in the herbs, atractyloside, which inhibits the ATP/ADP antiporter, was shown to inhibit oxidative phosphorylation of extramitochondrial ADP, but not intramitochondrial ADP. This finding demonstrated the importance of the ATP/ADP antiporter and has provided a powerful tool to study the mechanism by which this transporter functions.
Dioscorides lived near Tarsus, at the time a province of Rome in southeastern Asia Minor, in what is now Turkey. His five-volume De Materia Medica (The Materials of Medicine) “on the preparation, properties, and testing of drugs” described the medicinal properties of about 1000 natural products and 4740 medicinal usages of them. For approximately 1600 years, it was the basic reference in medicine from northern Europe to the Indian Ocean, comparable to today’s Physicians’ Desk Reference as a guide for using drugs.