Relative Activities of Photosystems I and II Are Regulated
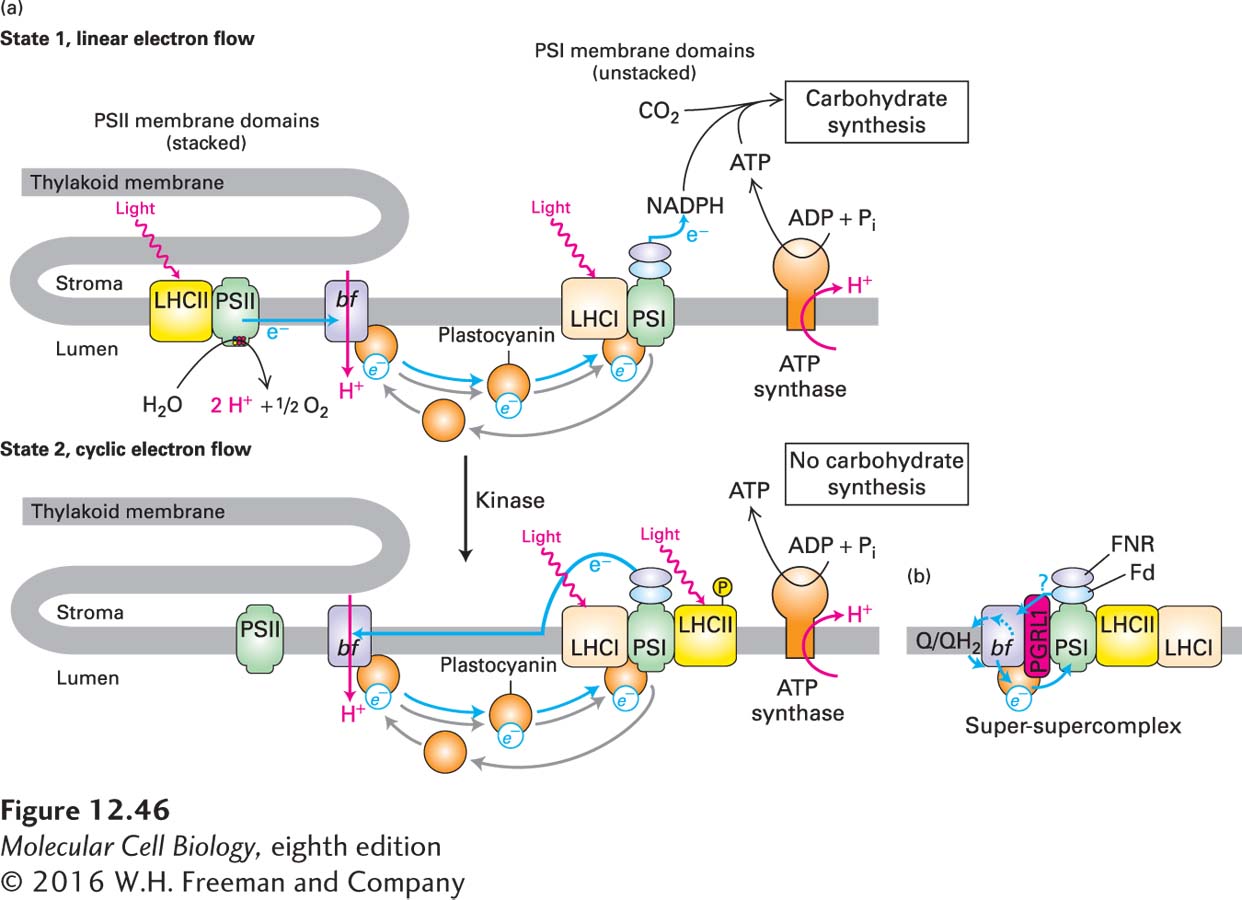
Chloroplasts respond to changes in the wavelengths and intensities of ambient light (as a consequence of the time of day, cloudiness, etc.) by altering the relative outputs of PSI and PSII to maintain the appropriate balance of ATP and NADPH production. In order for PSII, which is preferentially located in the stacked grana, and PSI, which is preferentially located in the unstacked thylakoid membranes, to act in sequence during linear electron flow, the amount of light energy delivered to the two reaction centers must be controlled so that each center activates the same number of electrons. This balanced condition is called state 1 (Figure 12-46a). If the two photosystems are not equally excited, then cyclic electron flow occurs in PSI, and PSII becomes less active (state 2).
Page 572
One mechanism underlying the regulation of the relative activities of PSI and PSII is the phosphorylation and dephosphorylation of thylakoid membrane proteins, including PSII and LHCII, mediated by several protein kinases and protein phosphatases. Changes in phosphorylation, particularly for LHCII, can alter the intra–
Under certain conditions, LHCII’s nonphosphorylated form is preferentially associated with PSII, and the phosphorylated form diffuses in the thylakoid membrane from the grana to the unstacked region and associates with PSI more than the nonphosphorylated form does. Light conditions in which there is preferential absorption of light by PSII result in activation of an LHCII kinase, increased LHCII phosphorylation, compensatory increased activation of PSI relative to PSII, and thus an increase in cyclic electron flow in state 2 (see Figure 12-46a). When the green alga Chlamydomonas reinhardtii was forced into state 2, it was possible to isolate a “super-
Regulating the supramolecular organization of the photosystems in plants has the effect of directing them toward ATP production (state 2) or toward the generation of reducing equivalents (NADPH) and ATP (state 1), depending on ambient light conditions and the metabolic needs of the plant. Both NADPH and ATP are required to convert CO2 to sucrose or starch, the fourth stage in photosynthesis, which we cover in the last section of this chapter.