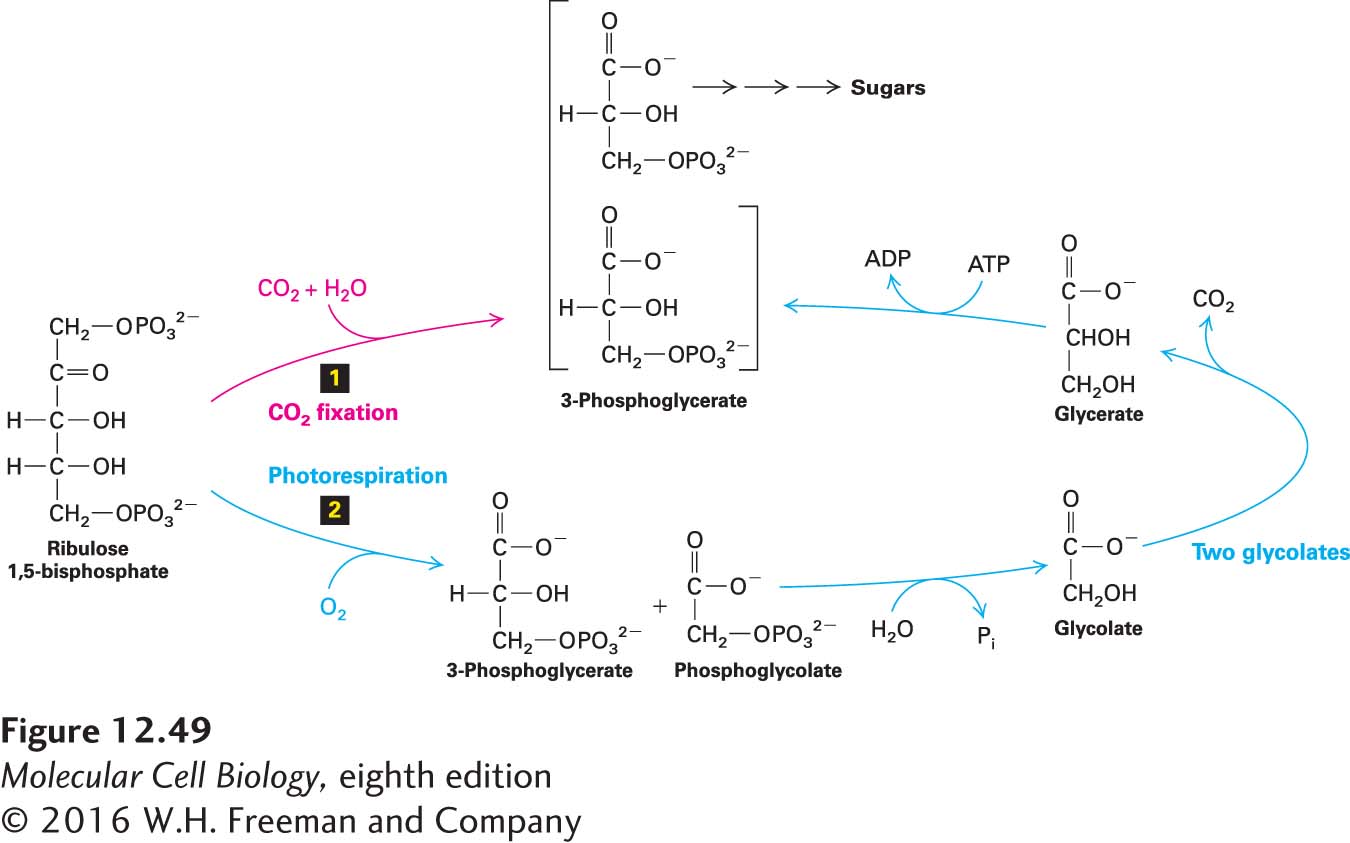
Photorespiration Competes with Carbon Fixation and Is Reduced in C4 Plants
As noted above, rubisco catalyzes the incorporation of CO2 into ribulose 1,5-bisphosphate as part of photosynthesis. It can catalyze a second, distinct, and competing reaction with the same substrate—ribulose 1,5-bisphosphate—but with O2 in place of CO2 as a second substrate, in a process known as photorespiration (Figure 12-49). The products of this second reaction are one molecule of 3-phosphoglycerate and one molecule of the two-carbon compound phosphoglycolate. The carbon-fixing reaction is favored when the ambient CO2 concentration is relatively high, whereas photorespiration is favored when CO2 is low and O2 is relatively high. Photorespiration takes place in light, consumes O2, and converts ribulose 1,5-bisphosphate in part to CO2. As Figure 12-49 shows, photorespiration is wasteful to the energy economy of the plant: it consumes ATP and O2, and it generates CO2 without fixing carbon. Indeed, when CO2 is low and O2 is high, much of the CO2 fixed by the Calvin cycle is lost as the result of photorespiration. This surprising, wasteful alternative reaction catalyzed by rubisco may be a consequence of the inherent difficulty the enzyme has in specifically binding the relatively featureless CO2 molecule and of the ability of both CO2 and O2 to react and form distinct products with the same initial enzyme/ribulose 1,5-bisphosphate intermediate.
FIGURE 12-49CO2 fixation and photorespiration. These competing pathways are both initiated by ribulose 1,5-bisphosphate carboxylase (rubisco), and both use ribulose 1,5-bisphosphate. CO2 fixation (pathway 1) is favored by high CO2 and low O2 concentrations; photorespiration (pathway 2) occurs at low CO2 and high O2 concentrations (that is, under normal atmospheric conditions). The phosphoglycolate produced by photorespiration is recycled via a complex set of reactions that take place in peroxisomes and in mitochondria, as well as in chloroplasts. The net result: for every two molecules of phosphoglycolate formed by photorespiration (four C atoms), one molecule of 3-phosphoglycerate is ultimately formed and recycled, and one molecule of CO2 is lost.
Excessive photorespiration could become a problem for plants in a hot, dry environment because they must keep the gas-exchange pores (stomata) in their leaves closed much of the time to prevent excessive loss of moisture. As a consequence, the CO2 level inside the leaf can fall below the Km of rubisco for CO2. Under these conditions, the rate of photosynthesis is slowed, photorespiration is greatly favored, and the plant might be in danger of fixing inadequate amounts of CO2. Corn, sugarcane, crabgrass, and other plants that can grow in hot, dry environments have evolved a way to avoid this problem by using a two-step pathway of CO2 fixation in which a CO2-hoarding step precedes the Calvin cycle. This pathway has been named the C4pathway because [14C]CO2 labeling showed that the first radioactive molecules formed during photosynthesis in this pathway are four-carbon compounds, such as oxaloacetate and malate, rather than the three-carbon molecules that initiate the Calvin cycle (C3 pathway).
Page 577
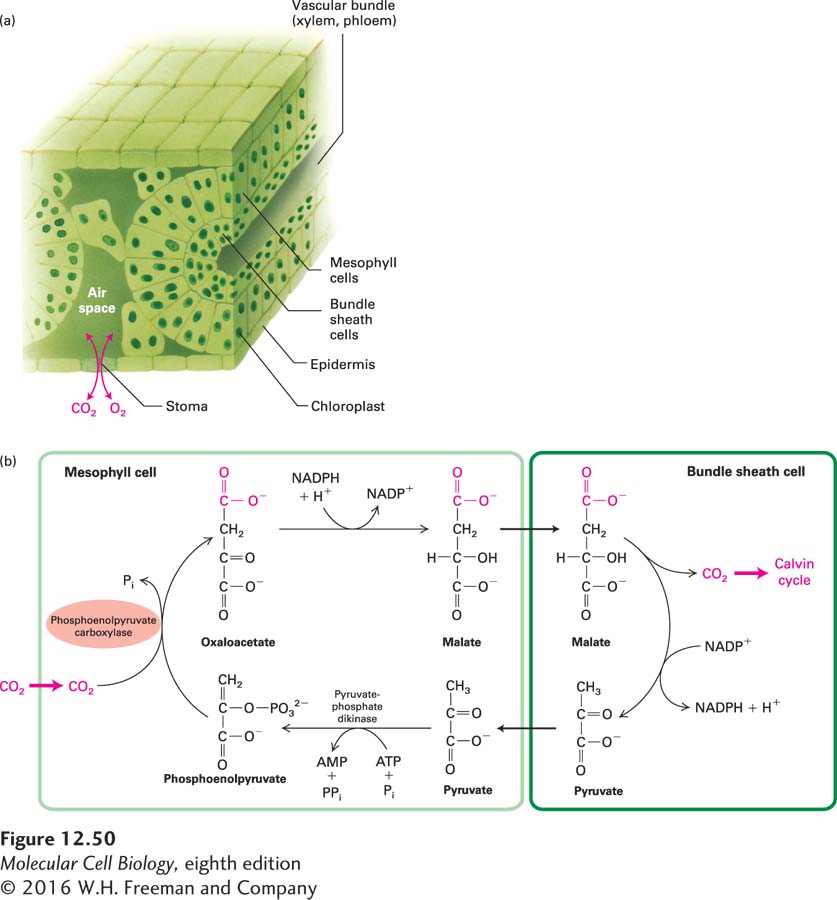
The C4 pathway involves two types of cells: mesophyll cells, which are adjacent to the air spaces in the leaf interior, and bundle sheath cells, which surround the vascular tissue and are sequestered away from the high oxygen levels to which mesophyll cells are exposed (Figure 12-50a). In the mesophyll cells of C4 plants, phosphoenolpyruvate, a three-carbon molecule derived from pyruvate, reacts with CO2 to generate oxaloacetate, a four-carbon compound (Figure 12-50b). The enzyme that catalyzes this reaction, phosphoenolpyruvate carboxylase, is found almost exclusively in C4 plants and, unlike rubisco, is insensitive to O2. The overall reaction that forms oxaloacetate from pyruvate involves the hydrolysis of one ATP and has a negative ΔG. Therefore, CO2 fixation will proceed even when the CO2 concentration is low. The oxaloacetate formed in the mesophyll cells is reduced to malate, which is transferred by a special transporter to the bundle sheath cells, where the CO2 is released by decarboxylation and enters the Calvin cycle (see Figure 12-50b).
FIGURE 12-50Leaf anatomy of C4 plants and the C4 pathway. (a) In C4 plants, bundle sheath cells line the vascular bundles containing the xylem and phloem. Mesophyll cells, which are adjacent to the substomal air spaces, can assimilate CO2 into four-carbon molecules at low ambient CO2 concentrations and deliver those molecules to the interior bundle sheath cells. The bundle sheath cells contain abundant chloroplasts and are the sites of photosynthesis and sucrose synthesis. Sucrose is carried to the rest of the plant via the phloem. In C3 plants, which lack bundle sheath cells, the Calvin cycle operates in the mesophyll cells to fix CO2. (b) The key enzyme in the C4 pathway is phosphoenolpyruvate carboxylase, which assimilates CO2 to form oxaloacetate in mesophyll cells. Decarboxylation of malate or other C4 intermediates in bundle sheath cells releases CO2, which enters the standard Calvin cycle (see Figure 12-48, top).
Because of the transport of CO2 from mesophyll cells, the CO2 concentration in the bundle sheath cells of C4 plants is much higher than it is in the normal atmosphere. Bundle sheath cells are also unusual in that they lack PSII and carry out only cyclic electron flow catalyzed by PSI, so no O2 is evolved. The high CO2 and reduced O2 concentrations in the bundle sheath cells favor the fixation of CO2 by rubisco to form 3-phosphoglycerate and suppress photorespiration.
Page 578
In contrast, the high O2 concentration in the atmosphere favors photorespiration in the mesophyll cells of C3 plants (pathway 2 in Figure 12-49); as a result, as much as 50 percent of the carbon fixed by rubisco may be reoxidized to CO2 in C3 plants. C4 plants are superior to C3 plants in their ability to use the available CO2 because the C4 enzyme phosphoenolpyruvate carboxylase has a higher affinity for CO2 than does rubisco. However, one ATP is converted to one AMP in the cyclical C4 process (to generate phosphoenolpyruvate from pyruvate); thus the overall efficiency of the photosynthetic production of sugars from NADPH and ATP is lower than it is in C3 plants, which use only the Calvin cycle for CO2 fixation. Nonetheless, the net rates of photosynthesis for C4 grasses, such as corn or sugarcane, can be two to three times the rates for otherwise similar C3 grasses, such as wheat, rice, or oats, owing to the elimination of losses from photorespiration.
Of the two carbohydrate products of photosynthesis, starch remains in the mesophyll cells of C3 plants and in the bundle sheath cells of C4 plants. In these cells, starch is subjected to glycolysis, mainly in the dark, forming ATP, NADH, and small molecules that are used as building blocks for the synthesis of amino acids, lipids, and other cellular constituents. Sucrose, in contrast, is exported from the photosynthetic cells and transported throughout the plant.