Mitochondria (Figure 12-6) are among the larger organelles in a eukaryotic cell. An individual mitochondrion is about the size of an E. coli bacterium, which is not surprising because, as we will see below, bacteria are thought to be the evolutionary precursors of mitochondria. Most eukaryotic cells contain many mitochondria, which may be fused to one another and may collectively occupy as much as 25 percent of the volume of the cytoplasm. The numbers of mitochondria in a cell—hundreds to thousands in mammalian cells—are regulated to match the cell’s requirements for ATP (e.g., specialized stomach cells that use a lot of ATP for acid secretion have many mitochondria).
[Part (b) T. G. Frey and G. A. Perkins. Part (c) from: Proc. Natl. Acad. Sci. USA 2013. 110 (22): 8936-8941, Fig. 6. Fig. 6A and C, “STED super-resolution microscopy reveals an array of MINOS clusters along human mitochondria,” by Jans et al.]
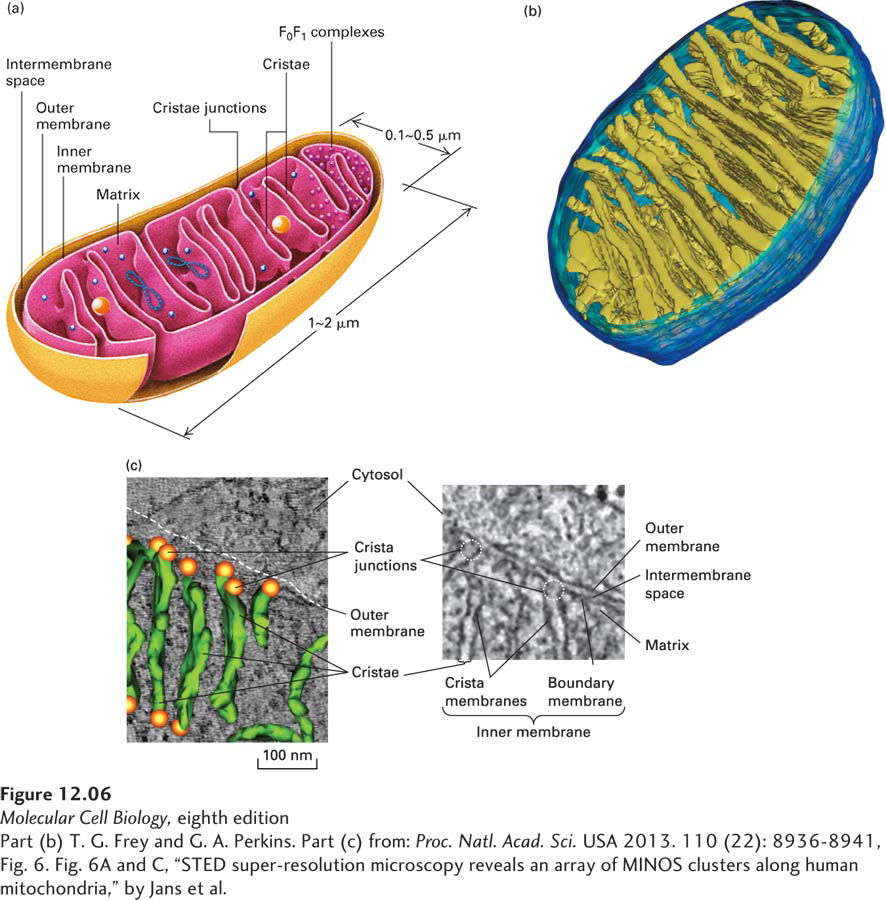
FIGURE 12-6Internal structure of a mitochondrion. (a) Schematic diagram showing the principal membranes and compartments. The smooth outer membrane forms the outside boundary of the mitochondrion. The inner membrane is apparently a single continuous membrane that has three distinct domains: boundary membrane, cristae, and crista junctions. The boundary membrane is flat and lies immediately below and adjacent to the outer membrane. The cristae are sheet-like and tubelike invaginations that extend from the boundary membrane into the center of the mitochondrion. The sharp bends that form the connection between the boundary membrane and the cristae are called crista junctions. The intermembrane space is continuous with the lumen of each crista. The F0F1 complexes (small red spheres), which synthesize ATP, are intramembrane particles that protrude from the cristae and inner membrane into the matrix. The matrix contains the mitochondrial DNA (blue strands), ribosomes (small blue spheres), and granules (large yellow spheres). (b) Computer-generated model of a section of a mitochondrion from chicken brain. This model is based on a three-dimensional electron microscopic image calculated from a series of two-dimensional electron micrographs recorded at regular intervals. This technique is analogous to a three-dimensional x-ray tomogram or CAT scan used in medical imaging. Note the tightly packed cristae (yellow-green), the inner membrane (light blue), and the outer membrane (dark blue). (c) Cristae and crista junctions from human fibroblasts were visualized and modeled using electron microscopy and tomography. The right panel shows one of the multiple sections through the mitochondrion imaged using transmission electron microscopy, with the mitochondrial membranes clearly distinguished. The sharp bends in the inner membrane at the junctions (dotted circles) that separate the crista membranes from the inner boundary membrane are seen clearly. The left panel shows a three-dimensional tomographic model of the laminar cristae seen edge on (green) and crista junctions (orange spheres) overlaid with the EM image.
[Part (b) T. G. Frey and G. A. Perkins. Part (c) from: Proc. Natl. Acad. Sci. USA 2013. 110 (22): 8936-8941, Fig. 6. Fig. 6A and C, “STED super-resolution microscopy reveals an array of MINOS clusters along human mitochondria,” by Jans et al.]
Although our focus in this chapter is on the mechanisms by which mitochondria generate ATP, it is important to recognize that mitochondria participate in a wide variety of critical cellular processes (Table 12-1). Mitochondria are involved with the biosynthesis of many small molecules. In several cases, some steps in these biosynthetic pathways take place within the mitochondria and some outside (extramitochondrial steps), requiring precursors and products to be shuttled into and out of the mitochondria via transport proteins. For example, in many eukaryotes, mitochondria host the first rate-limiting step in heme biosynthesis. The product of this first step is then exported for additional modification in the cytoplasm, producing a precursor that must then be imported into the mitochondria for the terminal steps. The biosynthesis in the cytoplasm of a wide variety of small molecules depends on mitochondria. Mitochondria provide to the cytoplasm small organic molecules—for example, citrate, isocitrate, malate, formate, and α-ketoglutarate—that can be used to generate NADPH (an energy source), acetyl CoA (a carbon source), or other precursors for the extramitochondrial biosynthesis of molecules such as glutathione, purines, fatty acids, and cholesterol.