A Preformed N-Linked Oligosaccharide Is Added to Many Proteins in the Rough ER
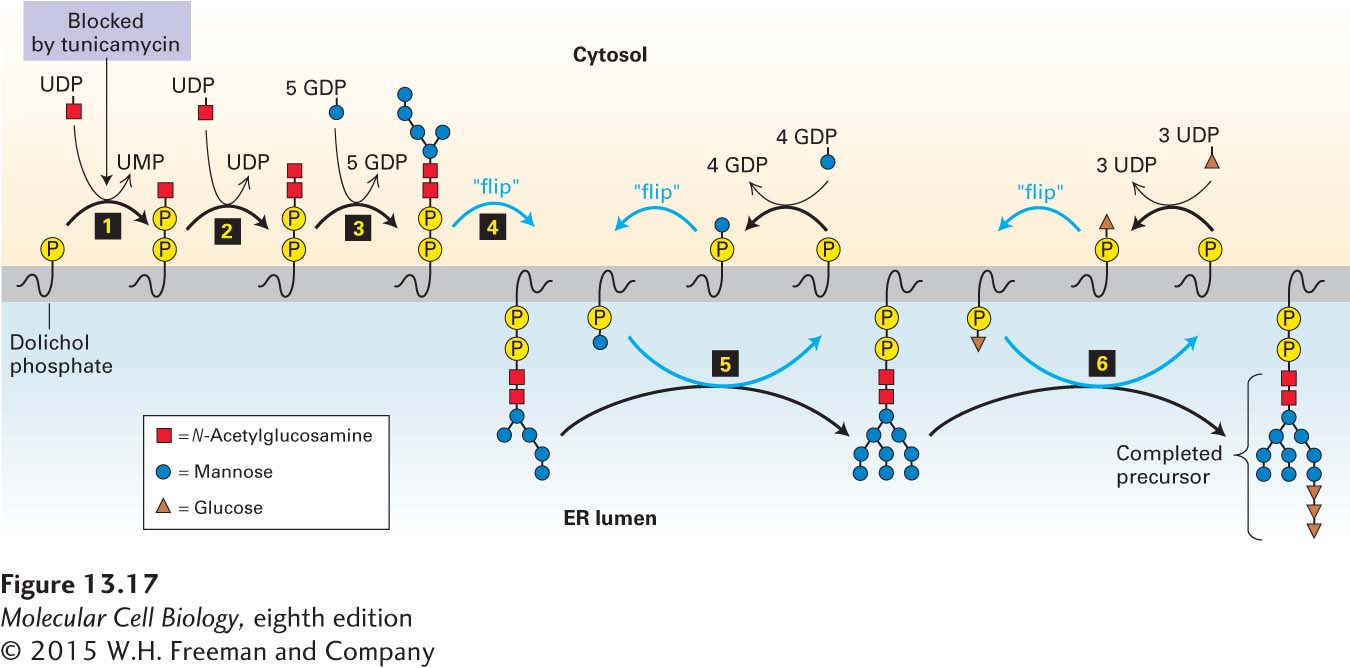
Biosynthesis of all N-linked oligosaccharides begins in the rough ER with a preformed oligosaccharide precursor containing 14 residues (Figure 13-17). The structure of this precursor is the same in plants, animals, and single-celled eukaryotes: a branched oligosaccharide, containing three glucose (Glc), nine mannose (Man), and two N-acetylglucosamine (GlcNAc) molecules, which can be written as Glc3Man9(GlcNAc)2. Once added to a protein, this branched carbohydrate structure is modified by addition or removal of monosaccharides in the ER and Golgi complex. The modifications to N-linked chains differ from one glycoprotein to another and among different organisms, but a core of 5 of the 14 residues is conserved in the structures of all N-linked oligosaccharides on secretory and membrane proteins.
FIGURE 13-17Biosynthesis of the oligosaccharide precursor. Dolichol phosphate is a strongly hydrophobic lipid, containing 75–95 carbon atoms, that is embedded in the ER membrane. Two N-acetylglucosamine (GlcNAc) and five mannose residues are added one at a time to a dolichol phosphate on the cytosolic face of the ER membrane (steps 1–3). The nucleotide-sugar donors in these and later reactions are synthesized in the cytosol. Note that the first sugar residue is attached to dolichol by a high-energy pyrophosphate linkage. Tunicamycin, which blocks the first enzyme in this pathway, inhibits the synthesis of all N-linked oligosaccharides in cells. After the seven-residue dolichol pyrophosphoryl intermediate is flipped to the luminal face (step 4), the remaining four mannose residues and all three glucose residues are added one at a time (steps 5–6). In the later reactions, the sugar to be added is first transferred from a nucleotide sugar to a carrier dolichol phosphate on the cytosolic face of the ER; the carrier is then flipped to the luminal face, where the sugar is transferred to the growing oligosaccharide, after which the “empty” carrier is flipped back to the cytosolic face. See C. Abeijon and C. B. Hirschberg, 1992, Trends Biochem. Sci.17:32.
Prior to transfer to a nascent chain in the lumen of the ER, the oligosaccharide precursor is assembled on a membrane-attached anchor called dolichol phosphate, a long-chain polyisoprenoid lipid (see Chapter 7). After the first sugar, GlcNAc, is attached to the dolichol phosphate by a pyrophosphate bond, the other sugars are added by glycosidic bonds in a complex set of reactions catalyzed by enzymes attached to the cytosolic or luminal faces of the rough ER membrane (see Figure 13-17). The final dolichol pyrophosphoryl oligosaccharide is oriented so that the oligosaccharide portion faces the ER lumen.
Page 602
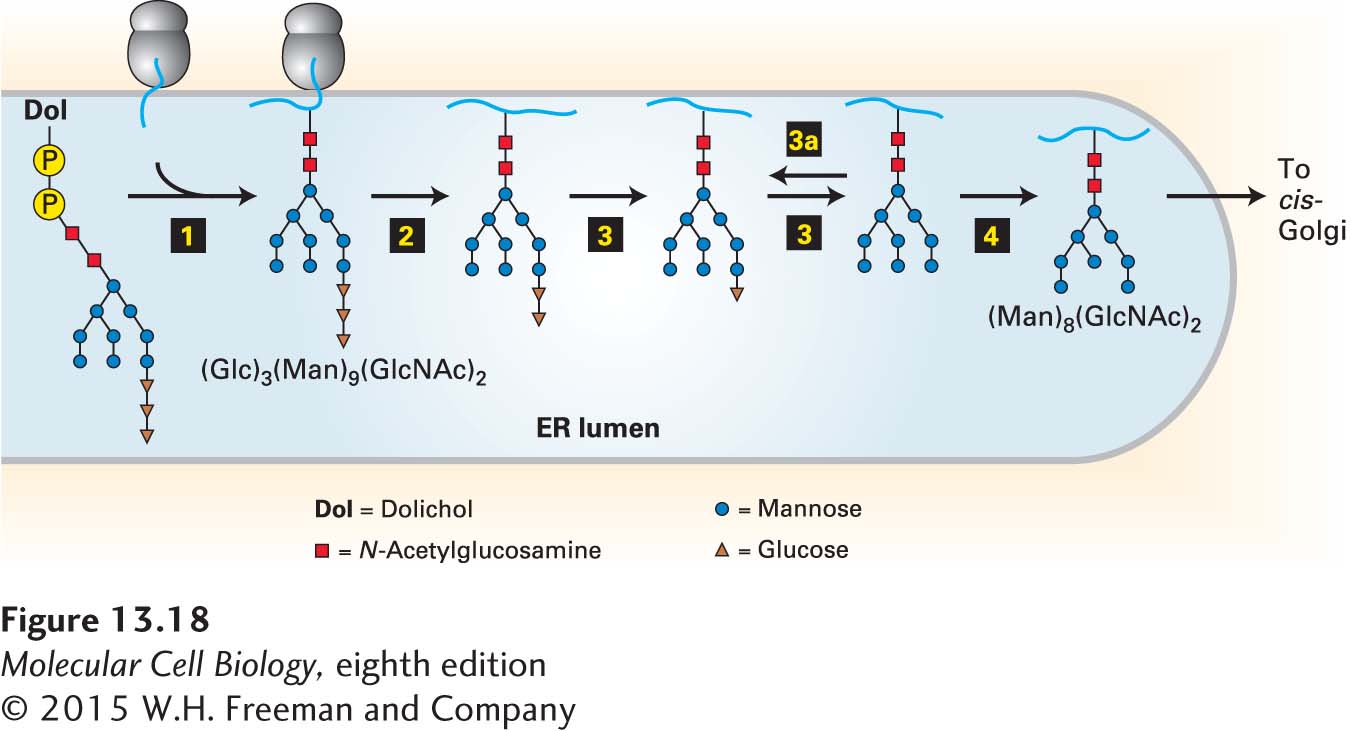
The entire 14-residue precursor is transferred from the dolichol carrier to an asparagine residue on a nascent polypeptide as it emerges into the ER lumen (Figure 13-18, step 1). Only asparagine residues in the tripeptide sequences Asn-X-Ser and Asn-X-Thr (where X is any amino acid except proline) are substrates for oligosaccharyl transferase, the enzyme that catalyzes this reaction. Two of the three subunits of this enzyme are ER membrane proteins whose cytosol-facing domains bind to the ribosome, localizing a third subunit of the transferase, the catalytic subunit, near the growing polypeptide chain in the ER lumen. Not all Asn-X-Ser/Thr sequences become glycosylated, and it is not possible to predict from the amino acid sequence alone which potential N-linked glycosylation sites will be modified; for instance, rapid folding of a segment of a protein containing an Asn-X-Ser/Thr sequence may prevent transfer of the oligosaccharide precursor to it.
FIGURE 13-18Addition and initial processing ofN-linked oligosaccharides. In the rough ER of vertebrate cells, the Glc3Man9(GlcNAc)2 precursor is transferred from the dolichol carrier to a susceptible asparagine residue on a nascent protein as soon as the asparagine crosses to the luminal side of the ER (step 1). In three separate reactions, first one glucose residue (step 2), then two glucose residues (step 3), and finally one mannose residue (step 4) are removed. Re-addition of one glucose residue (step 3a) plays a role in the correct folding of many proteins in the ER, as discussed later. The process of N-linked glycosylation of a soluble secretory protein is shown here, but the luminal portions of an integral membrane protein can be modified on asparagine residues by the same mechanism. See R. Kornfeld and S. Kornfeld, 1985, Annu. Rev. Biochem.45:631, and M. Sousa and A. J. Parodi, 1995, EMBO J.14:4196.
Immediately after the entire precursor, Glc3Man9(GlcNAc)2, is transferred to a nascent polypeptide, three different enzymes, called glycosidases, remove all three glucose residues and one particular mannose residue (Figure 13-18, steps 2–4). The three glucose residues, which are the last residues added during synthesis of the precursor on the dolichol carrier, appear to act as a signal that the oligosaccharide is complete and ready to be transferred to a protein.