A Cytosolic Receptor Targets Proteins with an SKL Sequence at the C-Terminus to the Peroxisomal Matrix
Peroxisomal-targeting sequences were first identified by testing of peroxisomal proteins with deletions for a specific defect in peroxisomal targeting. In one early study, the gene for firefly luciferase was expressed in cultured insect cells, and the resulting protein was shown to be properly targeted to the peroxisome. However, expression of a gene missing a small portion of the sequence encoding the C-terminus of the protein led to luciferase that failed to be targeted to the peroxisome and remained in the cytoplasm. By testing various mutant luciferase proteins in this system, researchers discovered that the sequence Ser-Lys-Leu (SKL in one-letter code) or a related sequence at the C-terminus is necessary for peroxisomal targeting. Furthermore, addition of the SKL sequence to the C-terminus of a normally cytosolic protein leads to uptake of the altered protein by peroxisomes in cultured cells. All but a few of the many different peroxisomal proteins bear a sequence of this type, known as peroxisomal-targeting sequence 1, or simply PTS1.
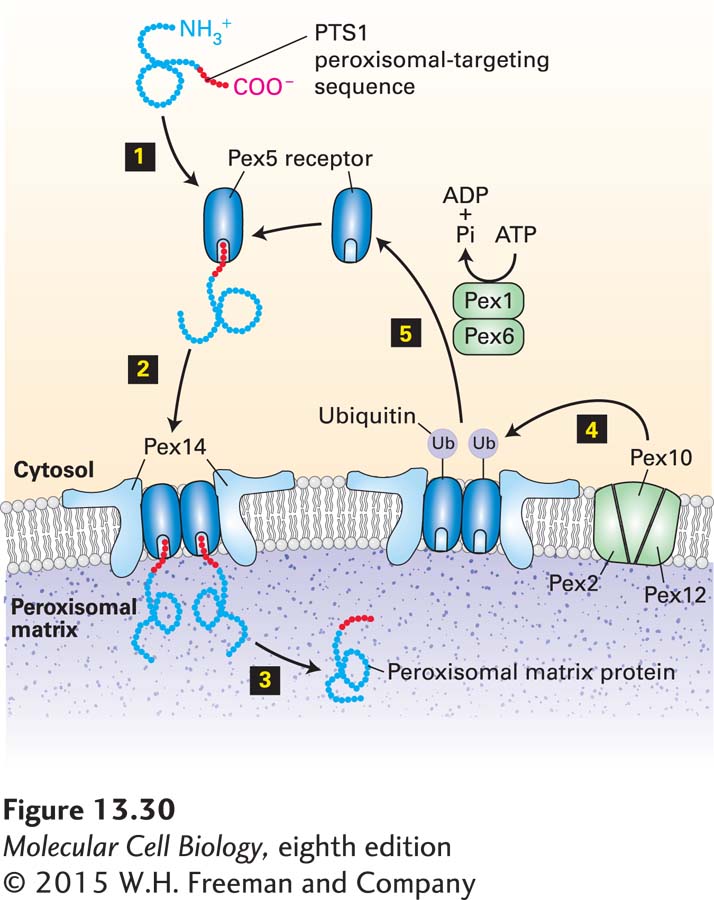
The pathway for the import of catalase and other PTS1-bearing proteins into the peroxisomal matrix is depicted in Figure 13-30. In the cytosol, PTS1 binds to a receptor called Pex5. Pex5 has the remarkable property of being able to switch from a monomeric soluble form to an oligomeric form embedded in the peroxisomal membrane in a complex with the membrane protein Pex14. In a manner that is not well understood, the PTS1-bearing protein is released from the oligomeric form of Pex5 into the interior of the peroxisome. This peroxisome import machinery, unlike most systems that mediate protein import into the ER, mitochondria, and chloroplasts, can translocate folded proteins across the membrane. For example, catalase assumes a folded conformation and binds to heme in the cytoplasm before traversing the peroxisomal membrane. Cell-free studies have shown that the peroxisome import machinery can transport large macromolecular objects, including gold particles about 9 nm in diameter, as long as they have a PTS1 tag attached to them. There is evidence that the size of oligomers of Pex5 bound to PTS1-bearing cargo molecules and Pex14 adjusts according to the size of the PTS1-bearing cargo molecules. The dynamic formation of oligomers is apparently the key mechanism by which PTS1-bearing cargo molecules can be accommodated without the formation of large stable pores that would disrupt the integrity of the peroxisomal membrane.
FIGURE 13-30PTS1-directed import of peroxisomal matrix proteins. Step 1: Most peroxisomal matrix proteins contain a C-terminal PTS1 targeting sequence (red), which binds to the cytosolic receptor Pex5. Step 2: Pex5 with the bound matrix protein forms a multimeric complex with the Pex14 receptor located on the peroxisomal membrane. Step 3: After assembly of the matrix protein-Pex5-Pex14 complex, the matrix protein dissociates from Pex5 and is released into the peroxisomal matrix. Steps 4 and 5: Pex5 is then returned to the cytosol by a process that involves ubiquitinylation by the membrane proteins Pex2, Pex10, and Pex12, followed by ATP-dependent removal from the membrane by the AAA-ATPase proteins Pex1 and Pex6. Note that folded proteins can be imported into peroxisomes and that the targeting sequence is not removed in the matrix. See P. E. Purdue and P. B. Lazarow, 2001, Annu. Rev. Cell Dev. Biol.17:701; S. Subramani et al., 2000, Annu. Rev. Biochem.69:399; and V. Dammai and S. Subramani, 2001, Cell105:187.
Page 620
Once the PTS1-bearing cargo molecule is released into the interior of the peroxisome, the oligomeric complex of Pex5 and Pex14 is actively disassembled, thus releasing Pex5 back into the cytoplasm in a soluble state. Pex5 recycling involves modification of membrane-bound Pex5 by ubiquitinylation. A complex of the peroxisomal membrane proteins Pex10, Pex12, and Pex2 transfers a ubiquitin moiety to Pex5. The AAA-ATPases Pex1 and Pex6, anchored to the peroxisomal membrane by Pex15, recognize ubiquitinylated Pex5 and use the energy from ATP hydrolysis to remove it from the oligomeric complex with Pex14 and release it into the cytosol. After removal of the ubiquitin modification, cytosolic Pex5 is ready to carry out another cycle of binding to a PTS1-bearing protein.
Peroxisomal import studies with purified components have shown that the binding of Pex5 to a PTS1-bearing protein, the assembly of an oligomeric complex of Pex5 and Pex14, and release of the PTS1-bearing protein into the interior of the peroxisome can all occur spontaneously without a source of chemical energy such as ATP. In contrast, both ubiquitin modification of Pex5 and the recycling of Pex5 by the AAA-ATPase are powered by ATP hydrolysis. Evidently, the recycling step in the import process uses energy to power unidirectional translocation of cargo molecules across the peroxisomal membrane.
Page 621
A few peroxisomal matrix proteins, such as thiolase, are synthesized as precursors with an N-terminal targeting sequence known as PTS2. These proteins bind to a different cytosolic receptor protein, but otherwise their import is thought to occur by the same mechanism as for PTS1-containing proteins.