Peroxisomal Membrane and Matrix Proteins Are Incorporated by Different Pathways
Autosomal recessive mutations that cause defective peroxisome assembly occur naturally in the human population. Such mutations can lead to severe developmental defects often associated with craniofacial abnormalities. In Zellweger syndrome and related disorders, for example, the transport of many or all proteins into the peroxisomal matrix is impaired: newly synthesized peroxisomal enzymes remain in the cytosol and are eventually degraded. Genetic analyses of cultured cells from patients with Zellweger syndrome and of yeast cells carrying similar mutations have identified more than 20 genes that are required for peroxisome biogenesis.
[Courtesy of Stephen Gould, Johns Hopkins University School of Medicine.]
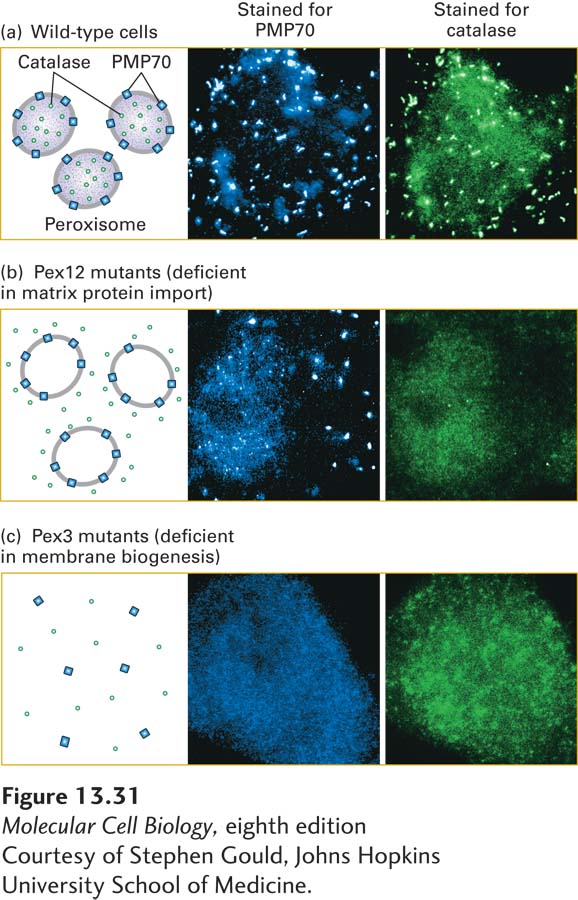
EXPERIMENTAL FIGURE 13-31Studies reveal different pathways for incorporation of peroxisomal membrane and matrix proteins. Cells were stained with fluorescent antibodies to PMP70, a peroxisomal membrane protein, or with fluorescent antibodies to catalase, a peroxisomal matrix protein, then viewed in a fluorescence microscope. (a) In wild-type cells, both peroxisomal membrane and matrix proteins are visible as bright foci in numerous peroxisomal bodies. (b) In cells from a Pex12-deficient patient, catalase is distributed uniformly throughout the cytosol, whereas PMP70 is localized normally to peroxisomal bodies. (c) In cells from a Pex3-deficient patient, peroxisomal membranes cannot assemble, and as a consequence, peroxisomal bodies do not form. Thus both catalase and PMP70 are mis-localized to the cytosol.
[Courtesy of Stephen Gould, Johns Hopkins University School of Medicine.]
Studies with peroxisome-assembly mutants have shown that different pathways are used for importing peroxisomal matrix proteins and for inserting proteins into the peroxisomal membrane (Figure 13-31). For example, analysis of cells from some patients with Zellweger syndrome led to the identification of the gene encoding Pex5 as well as many of the Pex genes needed for recycling of Pex5. Mutant cells that are defective in any one of these proteins cannot incorporate matrix proteins into peroxisomes; nonetheless, the cells contain empty peroxisomes that have a normal complement of peroxisomal membrane proteins (Figure 13-31b). Mutations in any one of three other genes were found to block insertion of peroxisomal membrane proteins as well as import of matrix proteins (Figure 13-31c). These findings demonstrate that one set of proteins translocates soluble proteins into the peroxisomal matrix, but a different set is required for insertion of proteins into the peroxisomal membrane. This situation differs markedly from that of the ER, mitochondrion, and chloroplast, whose membrane proteins and soluble proteins share many of the same components for their insertion into these organelles.
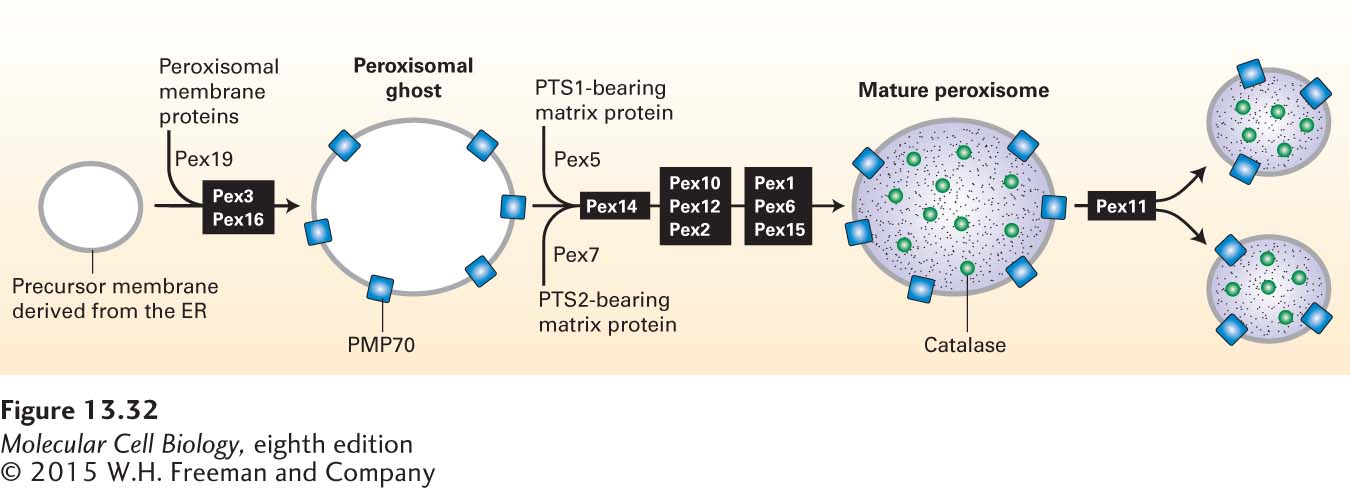
Although most peroxisomes are generated by the division of preexisting organelles, these organelles can arise de novo by the three-stage process depicted in Figure 13-32. In this case, peroxisome assembly begins in the ER. At least two peroxisomal membrane proteins, Pex3 and Pex16, are inserted into the ER membrane by the mechanisms described in Section 13.2. Pex3 and Pex16 then recruit Pex19 to form a specialized region of the ER membrane that can bud off of the ER to form a peroxisomal precursor membrane. Peroxisomal membrane protein assembly into mature peroxisomes also requires Pex19, which appears to act as a soluble receptor for peroxisomal membrane protein targeting sequences, as well as Pex3 and Pex16, which serve as membrane receptors for Pex19. The insertion of a complete set of peroxisomal membrane proteins produces membranes that have all the components necessary for import of matrix proteins, leading to the formation of mature, functional peroxisomes.
FIGURE 13-32Model of peroxisomal biogenesis and division. The first stage in the de novo formation of peroxisomes is the incorporation of peroxisomal membrane proteins into precursor membranes derived from the ER. Pex19 acts as the receptor for membrane-targeting sequences. A complex of Pex3 and Pex16 is required for proper insertion of proteins (e.g., PMP70) into the forming peroxisomal membrane. Insertion of all peroxisomal membrane proteins produces a peroxisomal ghost, which is capable of importing proteins targeted to the matrix. The pathways for importing PTS1- and PTS2-bearing matrix proteins differ only in the identity of the cytosolic receptor (Pex5 and Pex7, respectively) that binds the targeting sequence (see Figure 13-30). Complete incorporation of matrix proteins yields a mature peroxisome. Although peroxisomes can form de novo as just described, under most conditions, the proliferation of peroxisomes involves the division of mature peroxisomes, a process that depends on the Pex11 protein.
Division of mature peroxisomes, which largely determines the number of peroxisomes within a cell, depends on still another protein, Pex11. Overexpression of the Pex11 protein causes a large increase in the number of peroxisomes, suggesting that this protein controls the extent of peroxisome division. The small peroxisomes generated by division can be enlarged by incorporation of additional matrix and membrane proteins via the same pathways described previously.