Cotranslational Translocation Is Initiated by Two GTP-Hydrolyzing Proteins
Given that secretory proteins are synthesized in association with the ER membrane but not with any other cellular membrane, a signal-sequence recognition mechanism must target them there. The two key components in this targeting are the signal recognition particle (SRP) and its receptor. The SRP is a cytosolic ribonucleoprotein particle that transiently binds to both the ER signal sequence in a nascent protein and the large (60S) ribosomal subunit, forming a large complex. The SRP then targets the nascent protein–ribosome complex to the ER membrane by binding to the SRP receptor, which is located in the membrane.
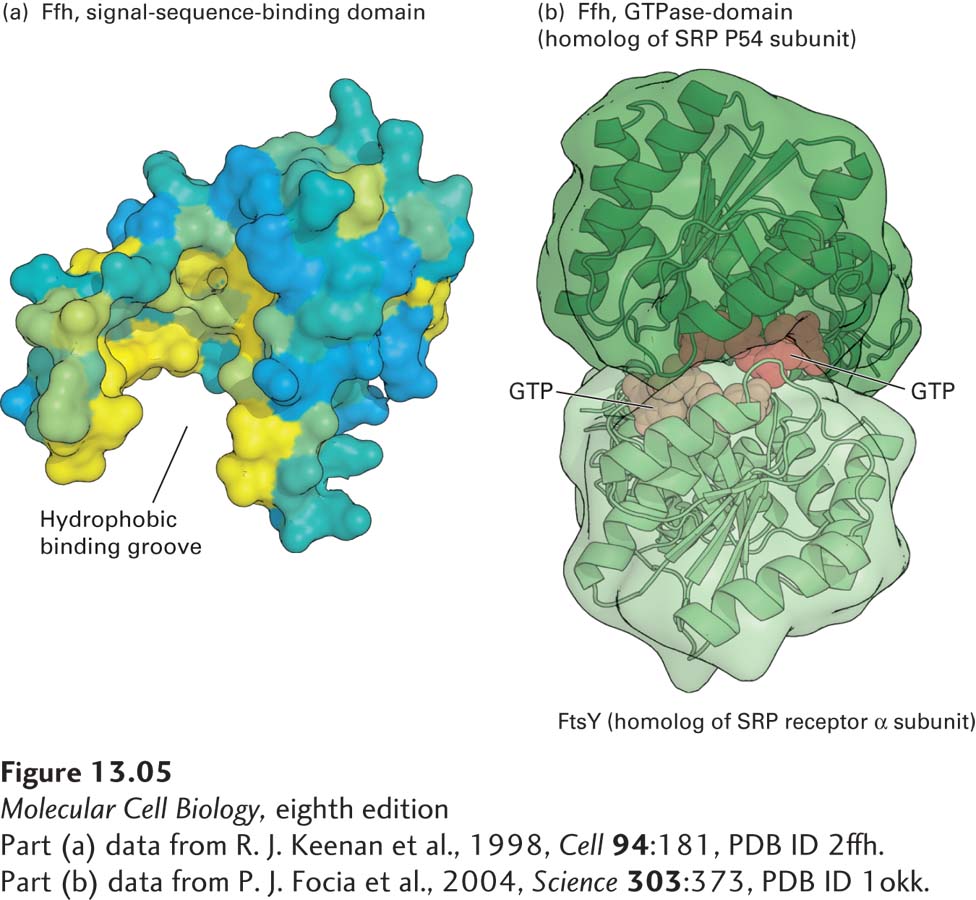
The SRP is made up of six proteins bound to a 300-nucleotide RNA, which acts as a scaffold for the hexamer. One of the SRP proteins (P54) can be chemically cross-linked to ER signal sequences, which shows that this subunit is the one that binds to the signal sequence in a nascent secretory protein. A region of P54 known as the M domain, containing many methionine and other amino acid residues with hydrophobic side chains, contains a cleft whose inner surface is lined by hydrophobic side chains (Figure 13-5a). The hydrophobic core of the signal sequence binds to this cleft via hydrophobic interactions. Other polypeptides in the SRP interact with the ribosome or are required for protein translocation into the ER lumen.
[Part (a) data from R. J. Keenan et al., 1998, Cell94:181, PDB ID 2ffh. Part (b) data from P. J. Focia et al., 2004, Science303:373, PDB ID 1okk.]
FIGURE 13-5Structure of the signal recognition particle (SRP). (a) The signal-sequence-binding domain: the bacterial Ffh protein is homologous to the portion of P54 that binds ER signal sequences in eukaryotes. This surface model shows the binding domain in Ffh, which contains a large cleft lined with hydrophobic amino acids (yellow) whose side chains interact with signal sequences. (b) GTP-and receptor-binding domain: the structure of GTP bound to FtsY (the archaeal homolog of the α subunit of the SRP receptor) and Ffh subunits from Thermus aquaticus illustrates how the interaction between these proteins is controlled by GTP binding and hydrolysis. Ffh and FtsY each can bind to one molecule of GTP, and when they bind to each other, the two bound molecules of GTP fit in the interface between the protein subunits and stabilize the dimer. Assembly of the pseudosymmetric dimer allows formation of two active sites for the hydrolysis of both bound GTP molecules. Hydrolysis to GDP destabilizes the interface, causing disassembly of the dimer.
[Part (a) data from R. J. Keenan et al., 1998, Cell94:181, PDB ID 2ffh. Part (b) data from P. J. Focia et al., 2004, Science303:373, PDB ID 1okk.]
Page 589
The SRP and the nascent polypeptide chain–ribosome complex bind to the ER membrane by docking with the SRP receptor, an integral protein of the ER membrane made up of two subunits: an α subunit and a smaller β subunit. Interaction of the SRP–nascent chain–ribosome complex with the SRP receptor is strengthened when both the P54 subunit of the SRP and the α subunit of the SRP receptor are bound to GTP. The structure of the SRP P54 subunit and the SRP receptor α subunit (FtsY) from the archaean Thermus aquaticus provides insight into how a cycle of GTP binding and hydrolysis can drive the binding and dissociation of these proteins. Figure 13-5b shows that P54 and FtsY, each bound to a single molecule of GTP, come together to form a pseudosymmetric heterodimer. Neither subunit alone contains a complete active site for the hydrolysis of GTP, but when the two proteins come together, they form two complete active sites that are capable of hydrolyzing both bound GTP molecules.
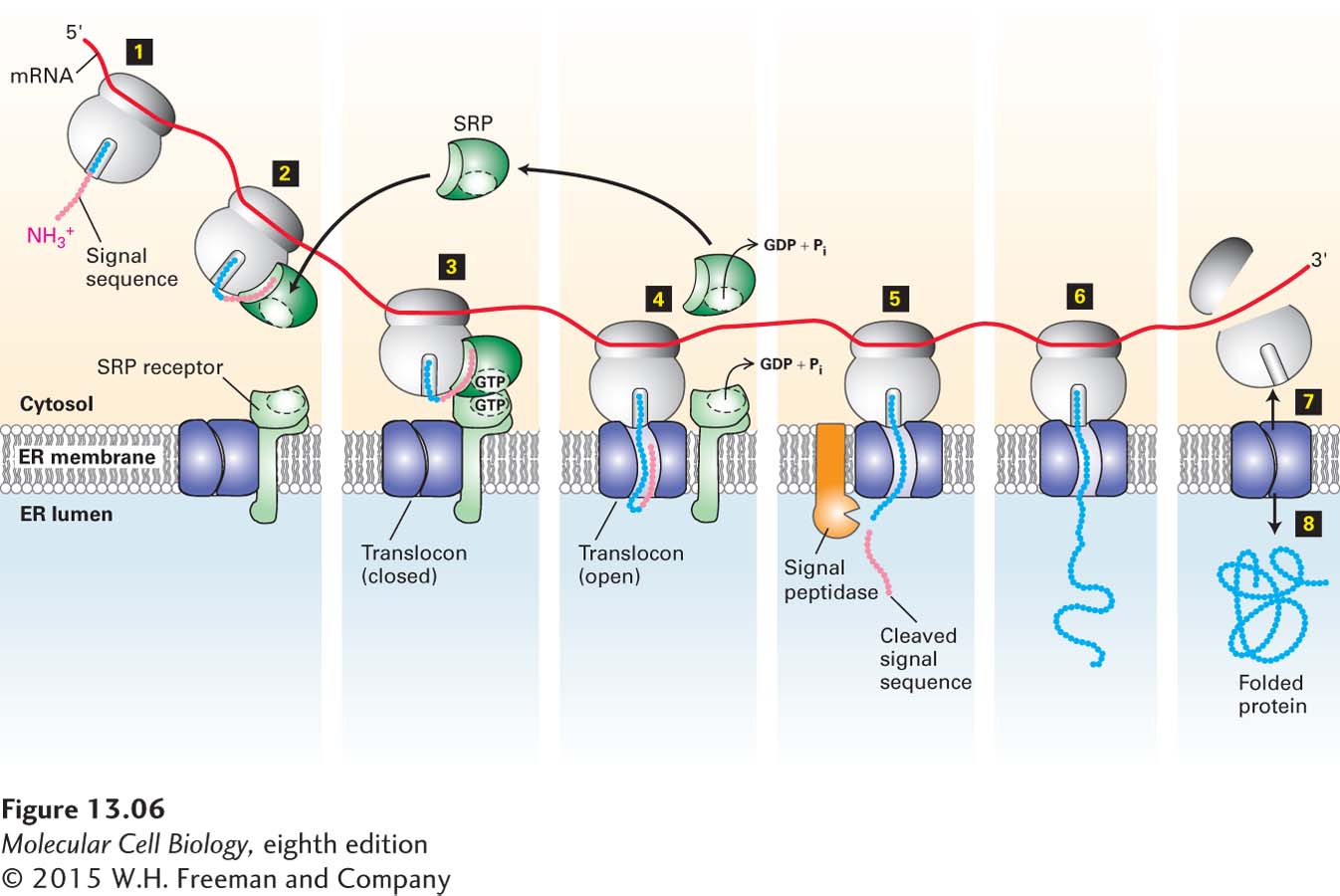
Figure 13-6 summarizes our current understanding of secretory protein synthesis and the role of the SRP and its receptor in this process. Hydrolysis of the bound GTP accompanies disassembly of the SRP and SRP receptor and initiates transfer of the nascent chain and ribosome to a site on the ER membrane, where translocation can take place. After dissociating from each other, the SRP and its receptor each release their bound GDP, SRP is recycled back to the cytosol, and both are ready to initiate another round of interaction between ribosomes synthesizing nascent secretory proteins and the ER membrane.
FIGURE 13-6Cotranslational translocation. Steps 1–2: Once the ER signal sequence emerges from the ribosome, it is bound by a signal recognition particle (SRP). Step 3: The SRP and the nascent polypeptide chain–ribosome complex bind to the SRP receptor in the ER membrane. This interaction is strengthened by the binding of GTP to both the SRP and its receptor. Step 4: Transfer of the nascent polypeptide–ribosome to the translocon leads to opening of this translocation channel to admit the growing polypeptide adjacent to the signal sequence, which is transferred to a hydrophobic binding site next to the central pore. Both the SRP and SRP receptor, once dissociated from the translocon, hydrolyze their bound GTP and then are ready to initiate the insertion of another polypeptide chain. Step 5: As the polypeptide chain elongates, it passes through the translocon channel into the ER lumen, where the signal sequence is cleaved by signal peptidase and is rapidly degraded. Step 6: The peptide chain continues to elongate as the mRNA is translated toward the 3′ end. Because the ribosome is attached to the translocon, the growing chain is extruded through the translocon into the ER lumen. Steps 7–8: Once translation is complete, the ribosome is released, the remainder of the protein is drawn into the ER lumen, the translocon closes, and the protein assumes its native folded conformation.