Passage of Growing Polypeptides Through the Translocon Is Driven by Translation
Once the SRP and its receptor have targeted a ribosome synthesizing a secretory protein to the ER membrane, the ribosome and nascent polypeptide chain are rapidly transferred to the translocon, a complex of proteins that forms a channel embedded within the ER membrane. As translation continues, the elongating chain passes directly from the large ribosomal subunit into the central pore of the translocon. The large ribosomal subunit is aligned with the pore of the translocon in such a way that the growing chain is never exposed to the cytoplasm and is prevented from folding until it reaches the ER lumen (see Figure 13-6).
Page 590
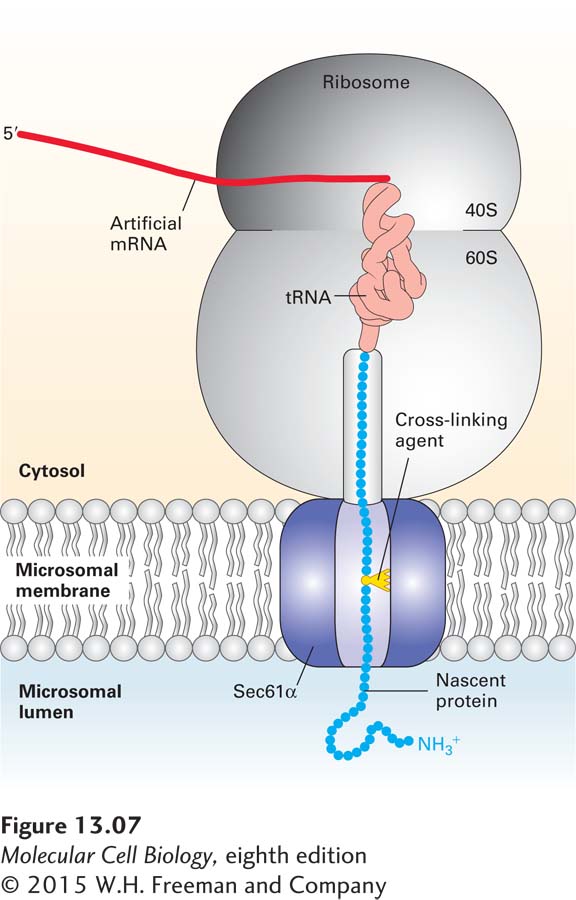
The translocon was first identified through mutations in the yeast gene encoding a protein called Sec61α, which caused a block in the translocation of secretory proteins into the lumen of the ER. Subsequently, three proteins, collectively called the Sec61 complex, were found to form the mammalian translocon: Sec61α, an integral membrane protein with 10 membrane-spanning α helices, and two smaller proteins, termed Sec61β and Sec61γ. Chemical cross-linking experiments in a cell-free translocation system—in which amino acid side chains from a nascent secretory protein became covalently attached to the Sec61α subunit—demonstrated that the translocating polypeptide chain comes into contact with the Sec61α protein, confirming its identity as the translocon pore (Figure 13-7).
FIGURE 13-7Sec61α is a translocon component. Cross-linking experiments show that Sec61 α is a translocon component that contacts nascent secretory proteins as they pass into the ER lumen. An mRNA encoding the N-terminal 70 amino acids of the secreted protein prolactin was translated in a cell-free system containing microsomes (see Figure 13-4b). The mRNA lacked a chain-termination codon and contained one lysine codon, near the middle of the sequence. The reaction mixtures contained a chemically modified lysyl-tRNA in which a light-activated cross-linking reagent was attached to the lysine side chain. Although the entire mRNA was translated, the completed polypeptide could not be released from the ribosome without a chain-termination codon and thus became “stuck” crossing the ER membrane. The reaction mixtures were then exposed to intense light, which caused the nascent polypeptide chain to become covalently bound to whatever proteins were near it in the translocon. When the experiment was performed using microsomes from mammalian cells, the nascent chain became covalently linked to Sec61α. Different versions of the prolactin mRNA were created so that the modified lysine residue would be placed at different distances from the ribosome; cross-linking to Sec61 α was observed only when the modified lysine was positioned within the translocation channel. See T. A. Rapoport, 1992, Science258:931 and D. Görlich and T. A. Rapoport, 1993, Cell75:615.
When microsomes in the cell-free translocation system were replaced with reconstituted phospholipid vesicles containing only the SRP receptor and the Sec61 complex, nascent secretory proteins were translocated from their SRP-ribosome complex into the vesicles. This finding indicates that the SRP receptor and the Sec61 complex are the only ER-membrane proteins that are absolutely required for translocation. Because neither of these proteins can hydrolyze ATP or otherwise provide energy to drive ongoing translocation, the energy derived from chain elongation at the ribosome appears to be sufficient to push the polypeptide chain across the membrane in one direction.
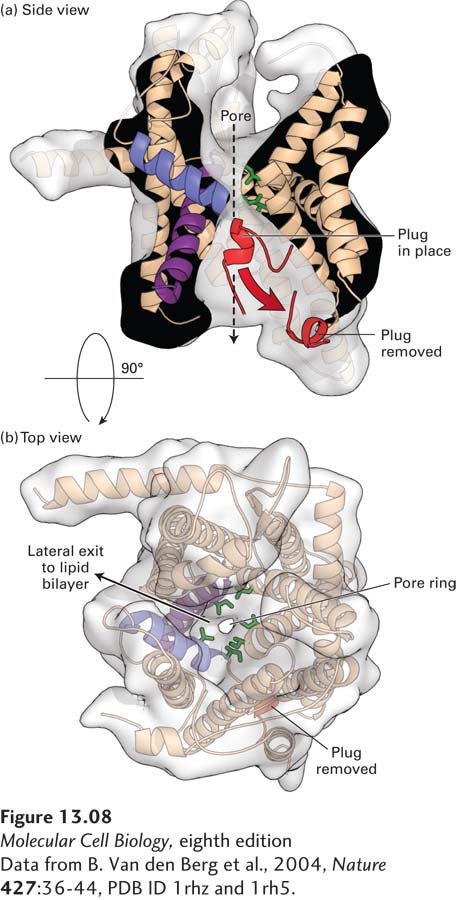
The translocon must be able to allow passage of a polypeptide chain while remaining sealed to small molecules, such as ATP, in order to maintain the permeability barrier of the ER membrane. Furthermore, there must be some way to regulate the translocon so that it is closed in its default state, opening only when a nascent polypeptide chain–ribosome complex is bound. A high-resolution structural model of the archaeal Sec61 complex shows how the translocon preserves the integrity of the membrane (Figure 13-8). The 10 transmembrane helices of Sec61α form a central channel through which the translocating polypeptide chain passes. Two different gating steps are required for Sec61α to accept a translocating polypeptide. The 10 transmembrane helices are organized into two 5-helix bundles. In the first gating step, the bundles hinge apart like an opening clamshell to expose a hydrophobic binding pocket for the hydrophobic core of the signal sequence at the open edge. The signal sequence binds to Sec61α with its N-terminus facing the cytosol and the elongating polypeptide doubling back through the central channel. The structural model of the Sec61 complex, which was isolated without a translocating peptide and is therefore presumed to be in a closed conformation, reveals a short helical peptide plugging the central channel. Biochemical studies of the Sec61 complex have shown that, in the absence of a translocating polypeptide, the peptide that forms the plug effectively seals the translocon to prevent the passage of ions and small molecules. In the second gating step, after the signal sequence has bound to the opened channel, the translocating peptide enters the central pore of the channel, forcing away the plug peptide and allowing translocation to proceed. The middle of the central pore is lined with hydrophobic isoleucine residues that in effect form a gasket, preventing leakage of small polar molecules around the translocating peptide even as translocation proceeds.
[Data from B. Van den Berg et al., 2004, Nature427:36–44, PDB ID 1rhz and 1rh5.]
FIGURE 13-8Structure of an archaeal Sec61 complex. The structure of the detergent-solubilized Sec61 complex from the archaeon M. jannaschii (also known as the SecY complex) was determined by x-ray crystallography. (a) A side view shows the hourglass-shaped channel through the center of the pore. A ring of isoleucine residues at the constricted waist of the pore forms a gasket that keeps the channel sealed to small molecules even as a translocating polypeptide passes through the channel. When no translocating peptide is present, the channel is closed by a short helical plug (red). This plug moves out of the channel during translocation. In this view, the front half of protein has been removed to better show the pore. (b) A view looking through the center of the channel shows a region (on the left side) where helices may separate, allowing lateral passage of a hydrophobic transmembrane domain into the lipid bilayer.
[Data from B. Van den Berg et al., 2004, Nature427:36–44, PDB ID 1rhz and 1rh5.]
Page 591
As the growing polypeptide chain enters the lumen of the ER, the signal sequence is cleaved by signal peptidase, which is a transmembrane ER protein associated with the translocon (see Figure 13-6, step 5). Signal peptidase recognizes a sequence on the C-terminal end of the hydrophobic core of the signal peptide and cleaves the chain specifically at this sequence once it has emerged into the luminal space of the ER. After the signal sequence has been cleaved, the growing polypeptide moves through the translocon into the ER lumen. The translocon remains open until translation is complete and the entire polypeptide chain has moved into the ER lumen. After translocation is complete, the plug peptide returns to the pore to reseal the translocon channel.