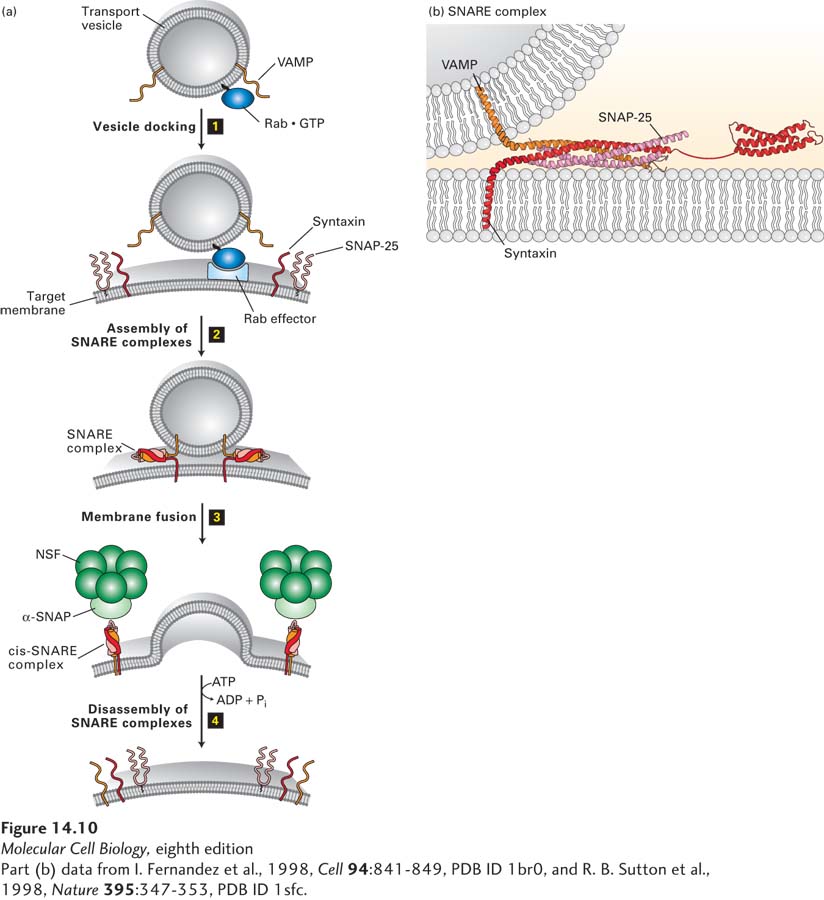
Rab GTPases Control Docking of Vesicles on Target Membranes
A second set of small GTP-binding proteins, known as Rab proteins, associate with transport vesicles and act as key regulators of vesicle trafficking to and fusion with the appropriate target membrane. Like Sar1 and ARF, Rab proteins belong to the GTPase superfamily of switch proteins. Rab proteins also contain an isoprenoid anchor that allows them to become tethered to the vesicle membrane. Association of an activated Rab protein with a specific vesicle type is generally a two-step process. In the first step, cytosolic Rab·GDP is targeted to the appropriate vesicle, becoming attached there by insertion of its isoprenoid anchor into the vesicle membrane. Often this attachment step is facilitated by a protein that can associate with Rab·GDP along with its isoprenoid anchor, usually known as a guanine nucleotide dissociation inhibitor (GDI). In the second step, a specific GEF located in the vesicle membrane converts membrane-bound Rab·GDP to Rab·GTP. Once localized and activated in this way, Rab·GTP is enabled to bind to a variety of different proteins, known as Rab effectors. Binding of Rab·GTP to a Rab effector can ultimately lead to docking of the vesicle on an appropriate target membrane (Figure 14-10a, step 1). After vesicle fusion occurs, the GTP bound to the Rab protein is hydrolyzed to GDP, triggering the release of Rab·GDP, which can then undergo another cycle of GDP-GTP exchange, binding, and hydrolysis.
[Part (b) data from I. Fernandez et al., 1998, Cell94:841-849, PDB ID 1br0, and R. B. Sutton et al., 1998, Nature395:347-353, PDB ID 1sfc.]
FIGURE 14-10Model for docking and fusion of transport vesicles with their target membranes. (a) The proteins shown in this example participate in fusion of secretory vesicles with the plasma membrane, but similar proteins mediate all vesicle-fusion events. Step 1: A Rab protein tethered via a lipid anchor to a secretory vesicle binds to an effector protein complex on the plasma membrane, thereby docking the transport vesicle on the appropriate target membrane. Step 2: A v-SNARE protein (in this case, VAMP) interacts with the cytosolic domains of the cognate t-SNAREs (in this case, syntaxin and SNAP-25). The very stable coiled-coil SNARE complexes that are formed hold the vesicle close to the target membrane. Step 3: Fusion of the two membranes immediately follows formation of SNARE complexes, but precisely how this occurs is not known. Step 4: Following membrane fusion, NSF, in conjunction with α-SNAP, binds to the SNARE complexes. The NSF-catalyzed hydrolysis of ATP then drives dissociation of the SNARE complexes, freeing the SNARE proteins for another round of vesicle fusion. Also at this time, Rab·GTP is hydrolyzed to Rab·GDP and dissociates from the Rab effector (not shown). (b) The SNARE complex. Numerous noncovalent interactions between four long α helices, two from SNAP-25 and one each from syntaxin and VAMP, stabilize the coiled-coil structure. See J. E. Rothman and T. Söllner, 1997, Science276:1212, Y. A. Chen and R. H. Scheller, 2001, Nat. Rev. Mol. Cell Biol. 2:98, and W. Weis and R. Scheller, 1998, Nature395:328.
[Part (b) data from I. Fernandez et al., 1998, Cell94:841-849, PDB ID 1br0, and R. B. Sutton et al., 1998, Nature395:347-353, PDB ID 1sfc.]
Page 642
A well-understood example of a Rab protein that enables vesicle fusion with the correct target membrane is the Sec4 protein of yeast, which specifically tags secretory vesicles, enabling them to fuse with the plasma membrane. Accordingly, yeast cells expressing mutant Sec4 proteins accumulate secretory vesicles that are unable to fuse with the plasma membrane (class E mutants in Figure 14-4). Sec4·GDP binds to secretory vesicles, where it is activated to Sec4·GTP by its cognate GEF, which is itself located on secretory vesicles. Sec4·GTP, in turn, binds to its effector, a large tethering complex composed of eight subunits, known as the exocyst. Tethering of secretory vesicles to the exocyst by binding of Sec4·GTP ultimately leads to vesicle fusion with the plasma membrane.
In mammalian cells, Rab5 protein is localized to endocytic vesicles, also known as early endosomes. These uncoated vesicles form from clathrin-coated vesicles just after they bud from the plasma membrane during endocytosis (see Figure 14-1, step 9). The fusion of early endosomes with one another in cell-free systems requires the presence of Rab5, and addition of Rab5 and GTP to cell-free extracts accelerates the rate at which these vesicles fuse with one another. A long coiled protein known as EEA1 (early endosome antigen 1), which resides on the membrane of the early endosome, functions as the effector for Rab5. In this case, Rab5·GTP on one endocytic vesicle is thought to bind specifically to EEA1 on the membrane of another endocytic vesicle, setting the stage for fusion of the two vesicles.
Other Rab proteins have motor proteins as effectors. For example, the Rab proteins Ypt31 and Ypt32, like Sec4, associate with secretory vesicles, but when in the activated GTP-bound state, recruit the effector myosin V to secretory vesicles. Myosin V enables secretory vesicles to move along actin filaments to the site of fusion with the plasma membrane.
Every type of transport vesicle appears to be labeled with one or more specific Rab proteins. These Rab proteins, through their specific association with effectors that are membrane tethers and molecular motors, ensure that the vesicles are directed to the correct target membrane address.