Anterograde Transport Through the Golgi Occurs by Cisternal Maturation
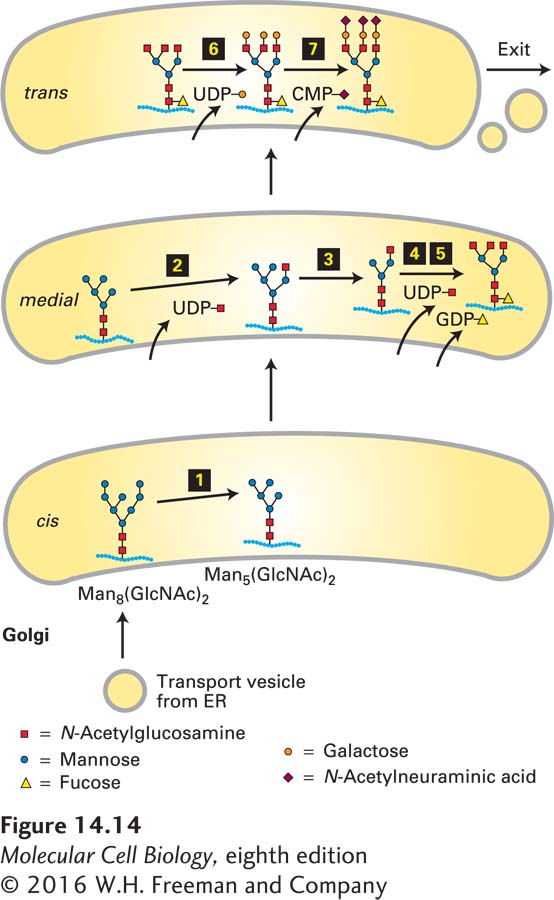
The Golgi complex is organized into three compartments, often arranged in a stacked set of flattened sacs, called cisternae. The compartments of the Golgi differ from one another according to the enzymes they contain. Many of the enzymes are glycosidases and glycosyltransferases that are involved in modifying the N-linked or O-linked carbohydrates attached to secretory proteins as they transit the Golgi complex. On the whole, the Golgi complex operates much like an assembly line, with proteins moving in sequence through the compartments, in which the modified carbohydrate chains in one compartment serve as the substrates for the modifying enzymes of the next compartment (Figure 14-14 shows a representative sequence of modification steps).
FIGURE 14-14Processing of N-linked oligosaccharide chains on glycoproteins within cis-, medial-, and trans-Golgi cisternae in vertebrate cells. The enzymes catalyzing each step are localized to the indicated compartments. After removal of three mannose residues in the cis-Golgi (step 1), the protein moves by cisternal maturation to the medial-Golgi. Here three N-acetylglucosamine (GlcNAc) residues are added (steps 2 and 4), two more mannose residues are removed (step 3), and a single fucose is added (step 5). Processing is completed in the trans-Golgi by addition of three galactose residues (step 6) and finally by linkage of an N-acetylneuraminic acid residue to each of the galactose residues (step 7). Specific transferase enzymes add sugars to the oligosaccharide, one at a time, from sugar nucleotide precursors imported from the cytosol. This pathway represents the Golgi processing events for a typical mammalian glycoprotein. Variations in the structure of N-linked oligosaccharides can result from differences in processing steps in the Golgi. See R. Kornfeld and S. Kornfeld, 1985, Annu. Rev. Biochem.45:631.
Page 649
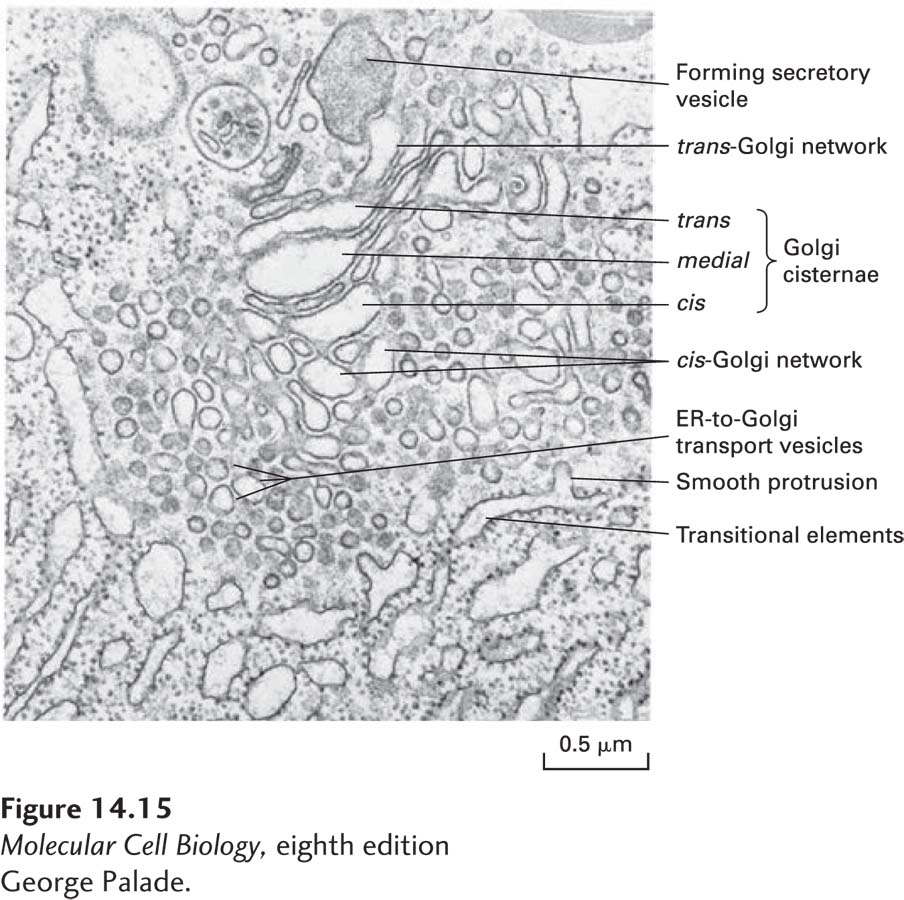
For many years, it was thought that the Golgi complex was an essentially static set of compartments with small transport vesicles carrying secretory proteins forward, from the cis- to the medial-Golgi and from the medial- to the trans-Golgi. Indeed, electron microscopy reveals many small vesicles associated with the Golgi complex that appear to move proteins from one Golgi compartment to another (Figure 14-15). However, these vesicles are now known to mediate retrograde transport, retrieving ER or Golgi enzymes from a later compartment and transporting them to an earlier compartment in the secretory pathway. Thus the Golgi appears to have a highly dynamic organization, continually forming transport vesicles, though only in the retrograde direction. To see the effect of this retrograde transport on the organization of the Golgi, consider the net effect on the medial-Golgi compartment as enzymes from the trans-Golgi move to the medial-Golgi while enzymes from the medial-Golgi are transported to the cis-Golgi. As this process continues, the medial-Golgi acquires enzymes from the trans-Golgi while losing medial-Golgi enzymes to the cis-Golgi and thus progressively becomes a new trans-Golgi compartment. In this way, secretory cargo proteins acquire carbohydrate modifications in the proper sequential order without being moved from one cisterna to another via anterograde vesicle transport.
[George Palade.]
EXPERIMENTAL FIGURE 14-15Electron micrograph of the Golgi complex in an pancreatic acinar cell reveals secretory and retrograde transport vesicles. A large secretory vesicle can be seen forming from the trans-Golgi network. Elements of the rough ER are on the bottom and left in this micrograph. Adjacent to the rough ER are transitional elements from which smooth protrusions appear to be budding. These buds form the small vesicles that transport secretory proteins from the rough ER to the Golgi complex. Interspersed among the Golgi cisternae are other small vesicles now known to function in retrograde, not anterograde, transport.
[George Palade.]
The first evidence that the forward transport of cargo proteins from the cis- to the trans-Golgi occurs by this progressive mechanism of cisternal maturation came from careful microscopic analysis of the synthesis of algal scales. These cell-wall glycoproteins are assembled in the cis-Golgi into large complexes visible in the electron microscope. Like other secretory proteins, newly made scales move from the cis- to the trans-Golgi, but they can be 20 times larger than the usual transport vesicles that bud from Golgi cisternae. Similarly, in the synthesis of collagen by fibroblasts, large aggregates of the procollagen precursor often form in the lumen of the cis-Golgi (see Figure 20-25). The procollagen aggregates are too large to be incorporated into small transport vesicles, and investigators could never find such aggregates in transport vesicles. These observations show that the forward movement of these, and perhaps all, secretory proteins from one Golgi compartment to another does not occur via small vesicles.
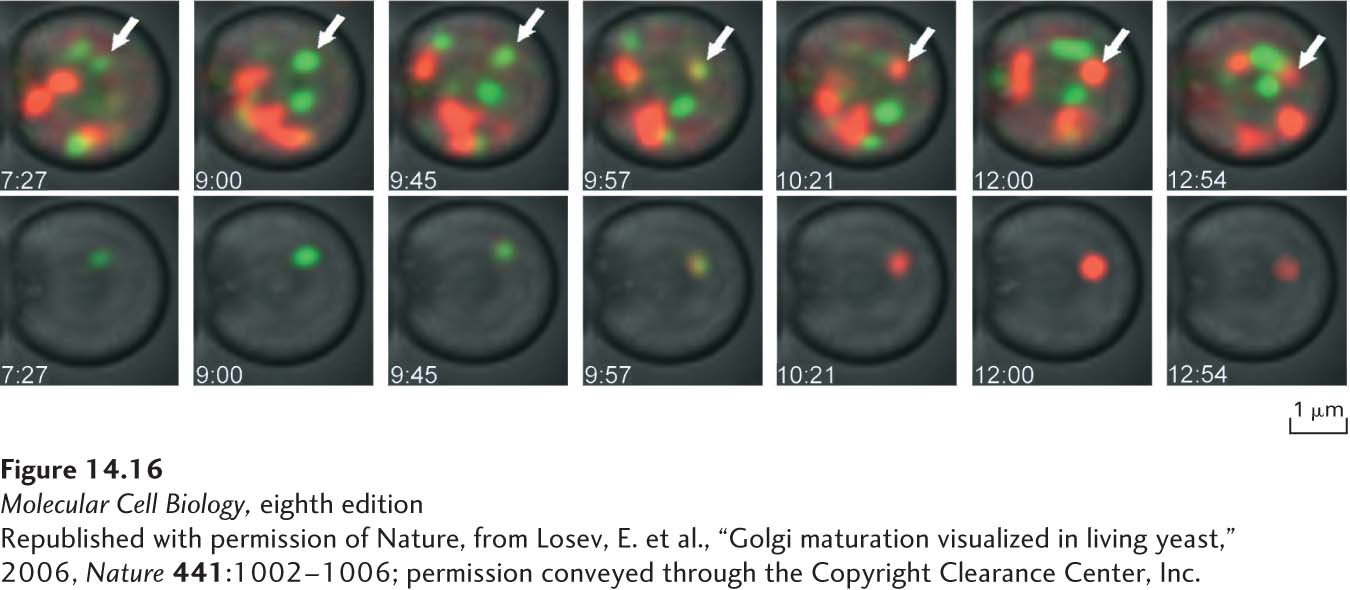
A particularly elegant demonstration of cisternal maturation in yeast takes advantage of different-colored fluorescent labels to image two different Golgi proteins simultaneously. Figure 14-16 shows how a cis-Golgi resident protein labeled with a green fluorescent protein and a trans-Golgi resident protein labeled with a red fluorescent protein behave in the same yeast cell. At any given moment, individual Golgi cisternae appear to have a distinct compartmental identity, in the sense that they contain either the cis-Golgi protein or the trans-Golgi protein, but only rarely contain both proteins. However, over time, an individual cisterna labeled with the cis-Golgi protein can be seen to progressively lose this protein and acquire the trans-Golgi protein. This behavior is exactly that predicted by the cisternal maturation model, in which the composition of an individual cisterna changes as Golgi resident proteins move from later to earlier Golgi compartments.
[Republished with permission of Nature, from Losev, E. et al., “Golgi maturation visualized in living yeast,” 2006, Nature441:1002–1006; permission conveyed through the Copyright Clearance Center, Inc.]
EXPERIMENTAL FIGURE 14-16Fluorescence-tagged fusion proteins demonstrate Golgi cisternal maturation in a live yeast cell. Yeast cells expressing the early Golgi protein Vrg4 fused to GFP (green fluorescence) and the late Golgi protein Sec7 fused to DsRed (red fluorescence) were imaged by time-lapse microscopy. The top series of images, taken approximately 1 minute apart, shows a collection of Golgi cisternae, which at any one time are labeled with either Vrg4 or Sec7. The bottom series of images show just one Golgi cisterna, isolated by digital processing of the image. First only Vrg4-GFP is located in the isolated cisterna, and later only Sec7-DsRed is located in the isolated cisterna, following a brief period in which both proteins are co-localized in this compartment. This experiment is a direct demonstration of the cisternal maturation hypothesis, showing that the composition of individual cisternae follows a process of maturation characterized by loss of early Golgi proteins and gain of late Golgi proteins.
[Republished with permission of Nature, from Losev, E. et al., “Golgi maturation visualized in living yeast,” 2006, Nature441:1002–1006; permission conveyed through the Copyright Clearance Center, Inc.]
Page 650
Although most protein traffic appears to move through the Golgi complex by a cisternal maturation mechanism, there is evidence that at least some of the COPI transport vesicles that bud from Golgi membranes contain cargo proteins (rather than Golgi enzymes) and move in an anterograde (rather than retrograde) direction.