Vesicles Coated with Clathrin and Adapter Proteins Mediate Transport from the trans-Golgi
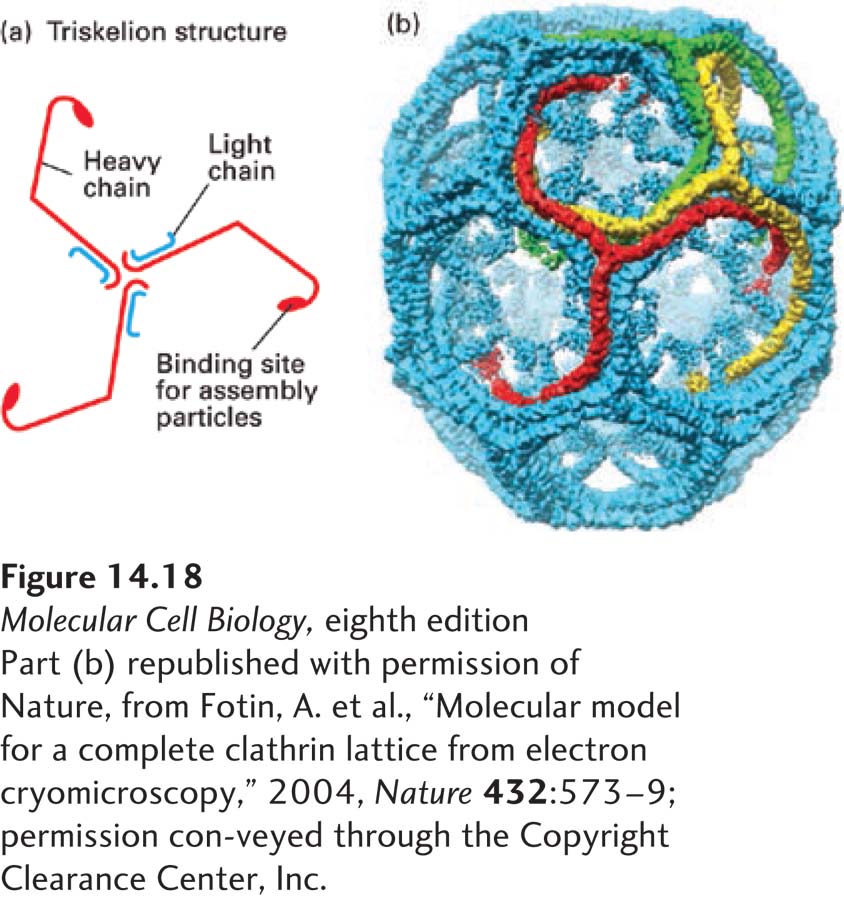
The best-characterized vesicles that bud from the trans-Golgi network have a two-layered coat: an outer layer composed of the fibrous protein clathrin and an inner layer composed of adapter protein (AP) complexes. Purified clathrin molecules, which have a three-limbed shape, are called triskelions, from the Greek for “three-legged” (Figure 14-18a). Each limb contains one clathrin heavy chain (180,000 MW) and one clathrin light chain (∼35,000–40,000 MW). Triskelions polymerize to form a polygonal lattice with an intrinsic curvature (Figure 14-18b). When clathrin polymerizes on a donor membrane, it does so in association with AP complexes, which fill the space between the clathrin lattice and the membrane. Each AP complex (340,000 MW) contains one copy each of four different adapter subunit proteins. A specific association between the globular domain at the end of each clathrin heavy chain in a triskelion and one subunit of the AP complex both promotes the co-assembly of clathrin triskelions with AP complexes and adds to the stability of the completed vesicle coat.
[Part (b) republished with permission of Nature, from Fotin, A. et al., “Molecular model for a complete clathrin lattice from electron cryomicroscopy,” 2004, Nature432:573–9; permission conveyed through the Copyright Clearance Center, Inc.]
FIGURE 14-18Structure of clathrin coats. (a) A clathrin molecule, called a triskelion, is composed of three heavy and three light chains. It has an intrinsic curvature due to the bend in the heavy chains. (b) Clathrin coats were formed in vitro by mixing purified clathrin heavy and light chains with AP2 complexes in the absence of membranes. Cryoelectron micrographs of more than 1000 assembled hexagonal clathrin barrel particles were analyzed by digital image processing to generate an average structural representation. The processed image shows only the clathrin heavy chains in a structure composed of 36 triskelions. Three representative triskelions are highlighted in red, yellow, and green. Some of the AP2 complexes packed into the interior of the clathrin cage are also visible in this representation. See B. Pishvaee and G. Payne, 1998, Cell95:443.
[Part (b) republished with permission of Nature, from Fotin, A. et al., “Molecular model for a complete clathrin lattice from electron cryomicroscopy,” 2004, Nature432:573–9; permission conveyed through the Copyright Clearance Center, Inc.]
By binding to the cytosolic face of membrane proteins, adapter proteins determine which cargo proteins are specifically included in (or excluded from) a budding transport vesicle. Three different AP complexes are known (AP1, AP2, AP3), each with four subunits of different, though related, proteins. Recently, a second general type of adapter protein, known as GGA, has been shown to contain in a single 70,000-MW polypeptide both clathrin- and cargo-binding elements similar to those found in the much larger hetero-tetrameric AP complexes. Vesicles containing each type of adapter complex (AP or GGA) have been found to mediate specific transport steps (see Table 14-1). All vesicles whose coats contain one of these complexes use ARF to initiate coat assembly on the donor membrane. As discussed previously, ARF also initiates assembly of COPI coats. The additional membrane features or protein factors that determine which type of coat will assemble after ARF attachment are not well understood at this time.
Vesicles that bud from the trans-Golgi network en route to the lysosome by way of the late endosome (see Figure 14-17, step 3) have clathrin coats associated with either AP1 or GGA. Both AP1 and GGA bind to the cytosolic domain of cargo proteins in the donor membrane. Membrane proteins containing a Tyr-X-X-Φ sequence, where X is any amino acid and Φ is a bulky hydrophobic amino acid, are recruited into clathrin/AP1-coated vesicles budding from the trans-Golgi network. This YXXΦsorting signal interacts with one of the AP1 subunits in the vesicle coat. As we discuss in the next section, vesicles with clathrin/AP2 coats, which bud from the plasma membrane during endocytosis, can also recognize the YXXΦ sorting signal. Vesicles coated with GGA proteins and clathrin bind cargo molecules with a different kind of sorting sequence. Cytosolic sorting signals that specifically bind to GGA adapter proteins include Asp-X-Leu-Leu and Asp-Phe-Gly-X-Φ sequences (where X and Φ are defined as above).
Page 652
Some vesicles that bud from the trans-Golgi network have coats composed of the AP3 complex. Although the AP3 complex does contain a binding site for clathrin similar to those in the AP1 and AP2 complexes, it is not clear whether clathrin is necessary for the functioning of AP3-containing vesicles because mutant versions of AP3 that lack the clathrin binding site appear to be fully functional. AP3-coated vesicles mediate trafficking to the lysosome, but they appear to bypass the late endosome and fuse directly with the lysosomal membrane (see Figure 14-17, step 2). In certain types of cells, such AP3 vesicles mediate protein transport to specialized storage compartments related to the lysosome. For example, AP3 is required for delivery of proteins to melanosomes, which contain the black pigment melanin in skin cells, and to platelet storage vesicles in megakaryocytes, large cells that fragment into dozens of platelets. Mice with mutations in either of two different subunits of AP3 not only have abnormal skin pigmentation but also exhibit bleeding disorders. The latter occur because platelets require normal storage vesicles in order to repair tears in blood vessels.