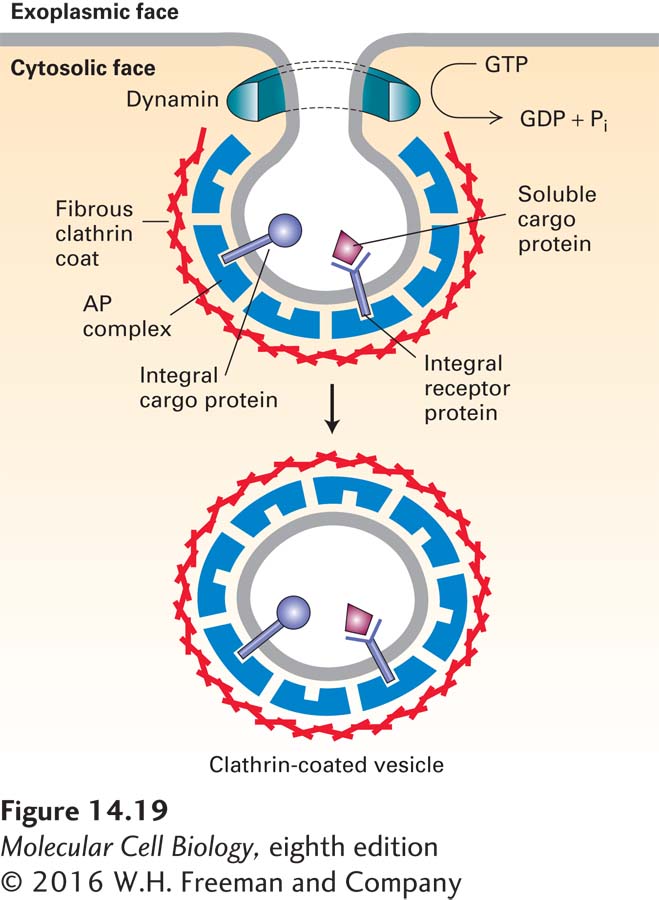
Dynamin Is Required for Pinching Off of Clathrin-Coated Vesicles
A fundamental step in the formation of a transport vesicle that we have not yet considered is how a vesicle bud is pinched off from the donor membrane. In the case of clathrin-coated vesicles, a cytosolic protein called dynamin is essential for the release of complete vesicles. At the later stages of bud formation, dynamin polymerizes around the neck portion of the bud and then hydrolyzes GTP. The energy derived from GTP hydrolysis is thought to drive a conformational change in dynamin that stretches the neck until the vesicle pinches off (Figure 14-19).
FIGURE 14-19Model for dynamin-mediated pinching off of clathrin-coated vesicles. After a vesicle bud forms, dynamin polymerizes over the neck. By a mechanism that is not well understood, dynamin-catalyzed hydrolysis of GTP leads to release of the vesicle from the donor membrane. Note that membrane proteins in the donor membrane are incorporated into vesicles by interacting with AP complexes in the coat. See K. Takel et al., 1995, Nature374:186.
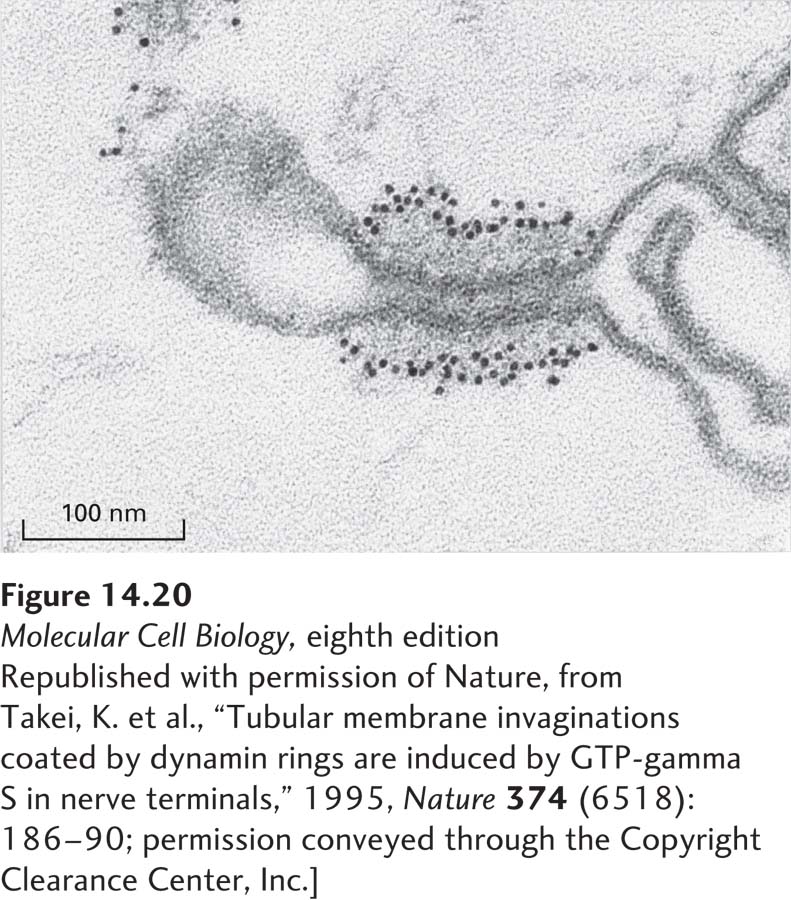
Incubation of cell extracts with a nonhydrolyzable derivative of GTP provides dramatic evidence for the importance of dynamin in the pinching off of clathrin/AP2-coated vesicles during endocytosis. Such treatment leads to accumulation of clathrin-coated vesicle buds with excessively long necks that are surrounded by polymeric dynamin but do not pinch off (Figure 14-20). Likewise, cells expressing mutant forms of dynamin that cannot bind GTP do not form clathrin-coated vesicles and instead accumulate similar long-necked vesicle buds encased with polymerized dynamin.
[Republished with permission of Nature, from Takei, K. et al., “Tubular membrane invaginations coated by dynamin rings are induced by GTP-gamma S in nerve terminals,” 1995, Nature374 (6518):186–90; permission conveyed through the Copyright Clearance Center, Inc.]
EXPERIMENTAL FIGURE 14-20GTP hydrolysis by dynamin is required for the pinching off of clathrin-coated vesicles in cell-free extracts. A preparation of nerve terminals, which undergo extensive endocytosis, was lysed by treatment with distilled water and incubated with GTP-γ-S, a nonhydrolyzable derivative of GTP. After sectioning, the preparation was treated with gold-tagged anti-dynamin antibody and viewed in the electron microscope. This image, which shows a long-necked clathrin/AP-coated bud with polymerized dynamin lining the neck, reveals that buds can form in the absence of GTP hydrolysis, but vesicles cannot pinch off. The extensive polymerization of dynamin that occurs in the presence of GTP-γ-S probably does not occur during the normal budding process.
[Republished with permission of Nature, from Takei, K. et al., “Tubular membrane invaginations coated by dynamin rings are induced by GTP-gamma S in nerve terminals,” 1995, Nature374 (6518):186–90; permission conveyed through the Copyright Clearance Center, Inc.]
Like COPI and COPII vesicles, clathrin-coated vesicles normally lose their coat soon after their formation. Cytosolic Hsp70, a constitutive chaperone protein found in all eukaryotic cells, is thought to use energy derived from the hydrolysis of ATP to drive depolymerization of the clathrin coat into triskelions. In the case of endocytic vesicles, uncoating not only releases triskelions for reuse in the formation of additional vesicles, but also exposes v-SNAREs for use in fusion with target membranes. Vesicle uncoating by cytosolic Hsp70 appears to be activated by a co-chaperone, auxillin, that contains a domain that stimulates the ATP hydrolysis by Hsp70. Conformational changes that occur when ARF switches from the GTP-bound to the GDP-bound state are thought to regulate the timing of clathrin coat depolymerization, but how the action of Hsp70 and auxillin is coupled to ARF switching is not well understood.