Some Proteins Undergo Proteolytic Processing After Leaving the trans-Golgi
For some secretory proteins (e.g., growth hormone) and certain viral membrane proteins (e.g., VSV G protein), removal of the N-terminal ER signal sequence from the nascent chain is the only known proteolytic cleavage required to convert the polypeptide to the mature, active protein (see Figure 13-6). However, some membrane proteins and many soluble secretory proteins are initially synthesized as relatively long-lived, inactive precursors, termed proproteins, that require further proteolytic processing to generate the mature, active proteins. Examples of proteins that undergo such processing are soluble lysosomal enzymes; many membrane proteins, such as influenza hemagglutinin (HA); and secreted proteins such as serum albumin, insulin, glucagon, and the yeast α mating factor. In general, the proteolytic conversion of a proprotein to the corresponding mature protein occurs after the proprotein has been sorted in the trans-Golgi network to appropriate vesicles.
In the case of soluble lysosomal enzymes, the proproteins, called proenzymes, are sorted by the M6P receptor as catalytically inactive enzymes. In the late endosome or lysosome, a proenzyme undergoes a proteolytic cleavage that generates a smaller but enzymatically active polypeptide. Delaying the activation of lysosomal proenzymes until they reach the lysosome prevents them from digesting macromolecules in earlier compartments of the secretory pathway.
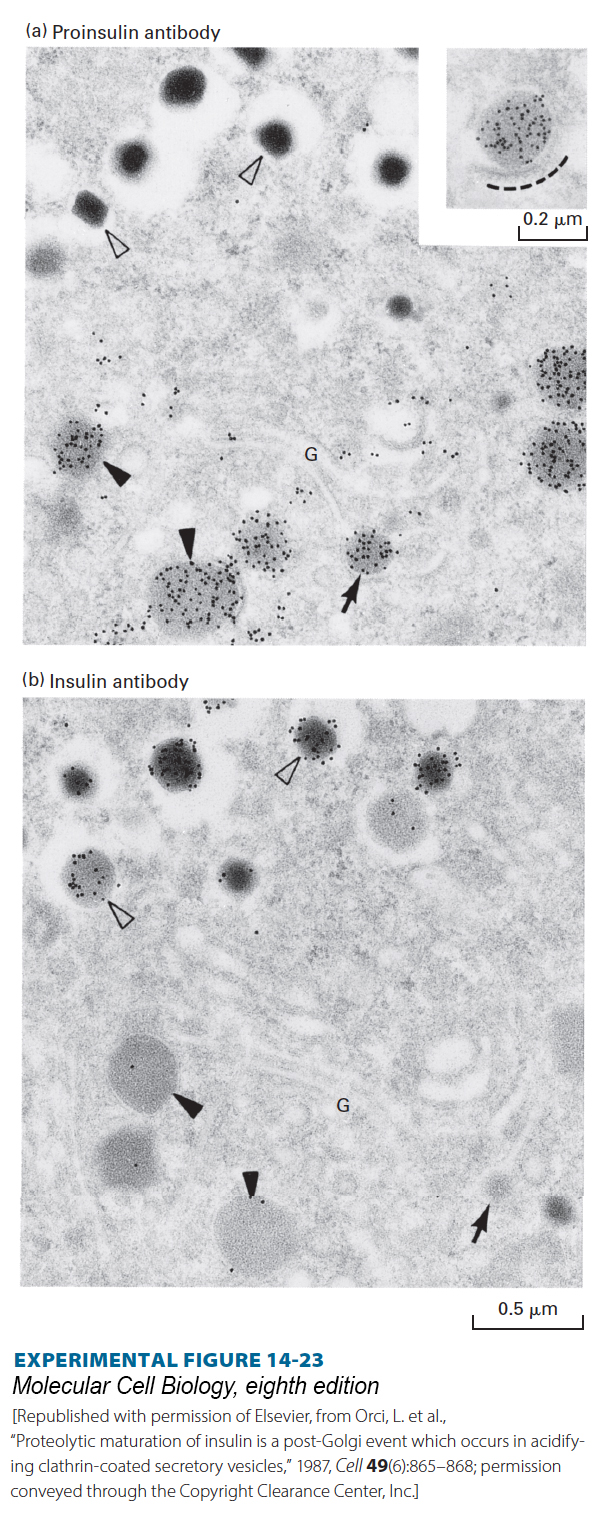
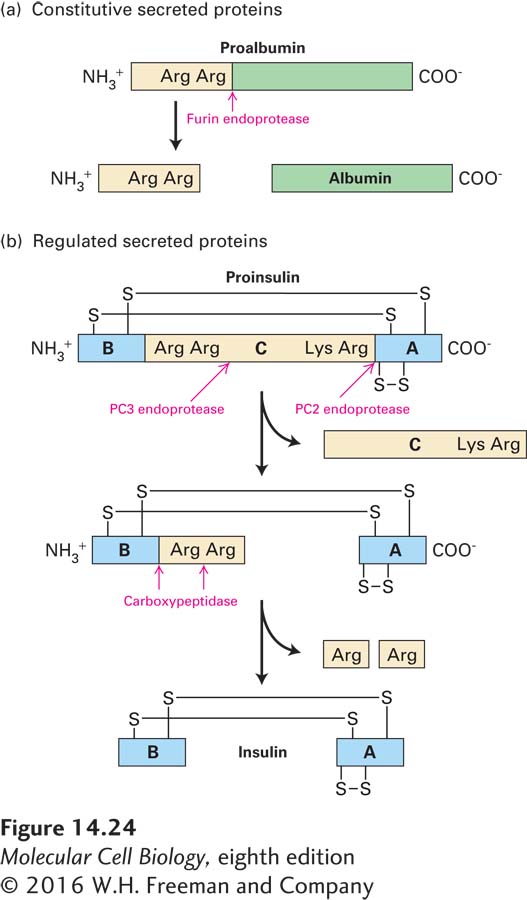
Normally, mature vesicles carrying secreted proteins to the cell surface are formed by fusion of several immature ones containing proprotein. Proteolytic cleavage of proproteins, such as proinsulin, occurs in vesicles after they move away from the trans-Golgi network (Figure 14-23). The proproteins of most constitutively secreted proteins (e.g., albumin) are cleaved only once at a site C-terminal to a dibasic recognition sequence such as Arg-Arg or Lys-Arg (Figure 14-24a). Proteolytic processing of proteins whose secretion is regulated generally entails additional cleavages. In the case of proinsulin, multiple cleavages of the single polypeptide chain yield the N-terminal B chain and the C-terminal A chain of mature insulin, which are linked by disulfide bonds, and the central C peptide, which is lost and subsequently degraded (Figure 14-24b).
[Republished with permission of Elsevier, from Orci, L. et al., “Proteolytic maturation of insulin is a post-Golgi event which occurs in acidifying clathrin-coated secretory vesicles,” 1987, Cell49(6):865–868; permission conveyed through the Copyright Clearance Center, Inc.]
EXPERIMENTAL FIGURE 14-23Proteolytic cleavage of proinsulin occurs in secretory vesicles after they have budded from the trans-Golgi network. Serial sections of the Golgi region of an insulin-secreting cell were stained with (a) a monoclonal antibody that recognizes proinsulin, but not insulin, or (b) a different antibody that recognizes insulin, but not proinsulin. The antibodies, which were bound to electron-opaque gold particles, appear as dark dots in these electron micrographs (see Figure 4-33). Immature secretory vesicles (closed arrowheads) and vesicles budding from the trans-Golgi (arrows) stain with the proinsulin antibody, but not with the insulin antibody. These vesicles contain diffuse protein aggregates that include proinsulin and other regulated secreted proteins. Mature vesicles (open arrowheads) stain with insulin antibody, but not with proinsulin antibody, and have a dense core of almost crystalline insulin. Since budding and immature secretory vesicles contain proinsulin (not insulin), the proteolytic conversion of proinsulin to insulin must take place in these vesicles after they bud from the trans-Golgi network. The inset in (a) shows a proinsulin-rich secretory vesicle surrounded by a protein coat (dashed line).
[Republished with permission of Elsevier, from Orci, L. et al., “Proteolytic maturation of insulin is a post-Golgi event which occurs in acidifying clathrin-coated secretory vesicles,” 1987, Cell49(6):865–868; permission conveyed through the Copyright Clearance Center, Inc.]
FIGURE 14-24Proteolytic processing of proproteins in the constitutive and regulated secretory pathways. The processing of proalbumin and proinsulin is typical of the constitutive and regulated pathways, respectively. The endoproteases that function in such processing cleave at the C-terminal end of a sequence of two consecutive amino acids. (a) The endoprotease furin acts on the precursors of constitutive secreted proteins. (b) Two endoproteases, PC2 and PC3, act on the precursors of regulated secreted proteins. The final processing of many such proteins is catalyzed by a carboxypeptidase that sequentially removes two basic amino acid residues at the C-terminus of a polypeptide. See D. Steiner et al., 1992, J. Biol. Chem.267:23435.
Page 657
The breakthrough in identifying the proteases responsible for such processing of secreted proteins came from analysis of yeast with a mutation in the KEX2 gene. These mutant cells synthesized the precursor of the α mating factor but could not proteolytically process it to the functional form and thus were unable to mate with cells of the opposite mating type. The wild-type KEX2 gene encodes an endoprotease that cleaves the α-factor precursor at a site C-terminal to Arg-Arg and Lys-Arg residues. Mammals contain a family of endoproteases homologous to the yeast KEX2 protein, all of which cleave a protein chain on the C-terminal side of an Arg-Arg or Lys-Arg sequence. One, called furin, is found in all mammalian cells; it processes proteins such as albumin that are secreted constitutively. In contrast, the PC2 and PC3 endoproteases are found only in cells that exhibit regulated secretion; these enzymes are localized to regulated secretory vesicles and proteolytically cleave the precursors of many hormones at specific sites.