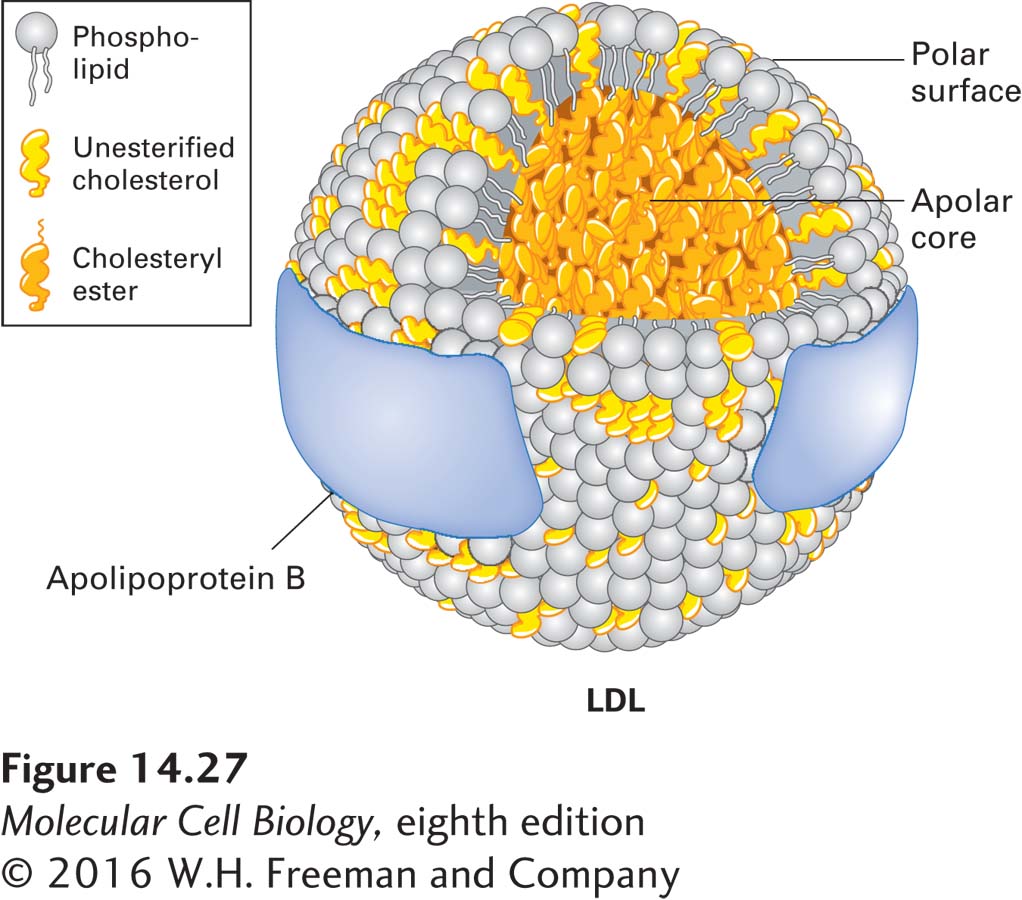
Cells Take Up Lipids from the Blood in the Form of Large, Well-Defined Lipoprotein Complexes
Lipids absorbed from the diet in the intestines or stored in adipose tissue can be distributed to cells throughout the body. To facilitate the mass transfer of lipids between cells, animals have evolved an efficient way to package from hundreds to thousands of lipid molecules into water-soluble, macromolecular carriers, called lipoproteins, that cells can take up from the circulation as an ensemble. A lipoprotein particle has a shell composed of proteins (apolipoproteins) overlying a cholesterol-containing phospholipid monolayer. The shell is amphipathic because its outer surface is hydrophilic, making the particle water soluble, and its inner surface is hydrophobic. Beneath the hydrophobic inner surface of the shell is a core of neutral lipids containing mostly cholesteryl esters, triglycerides, or both. Mammalian lipoproteins fall into different classes, defined by their differing buoyant densities. The class we will consider here is low-density lipoprotein (LDL). A typical LDL particle, depicted in Figure 14-27, is a sphere 20–25 nm in diameter. The amphipathic outer shell is composed of a phospholipid monolayer and a single molecule of a large protein known as apoB-100; the core of the particle is packed with cholesterol in the form of cholesteryl esters.
FIGURE 14-27Model of low-density lipoprotein (LDL). All classes of lipoproteins have the same general structure: an amphipathic shell composed of apolipoprotein, a phospholipid monolayer (not bilayer), and cholesterol, and a hydrophobic core composed mostly of cholesteryl esters or triglycerides, or both, but with minor amounts of other neutral lipids (e.g., some vitamins). This model of LDL is based on electron microscopy and other low-resolution biophysical methods. LDL is unique in that it contains only a single molecule of one type of apolipoprotein (ApoB), which appears to wrap around the outside of the particle as a band of protein. The other lipoproteins contain multiple apolipoprotein molecules, often of different types. See M. Krieger, 1995, in E. Haber, ed., Molecular Cardiovascular Medicine, Scientific American Medicine, pp. 31–47.
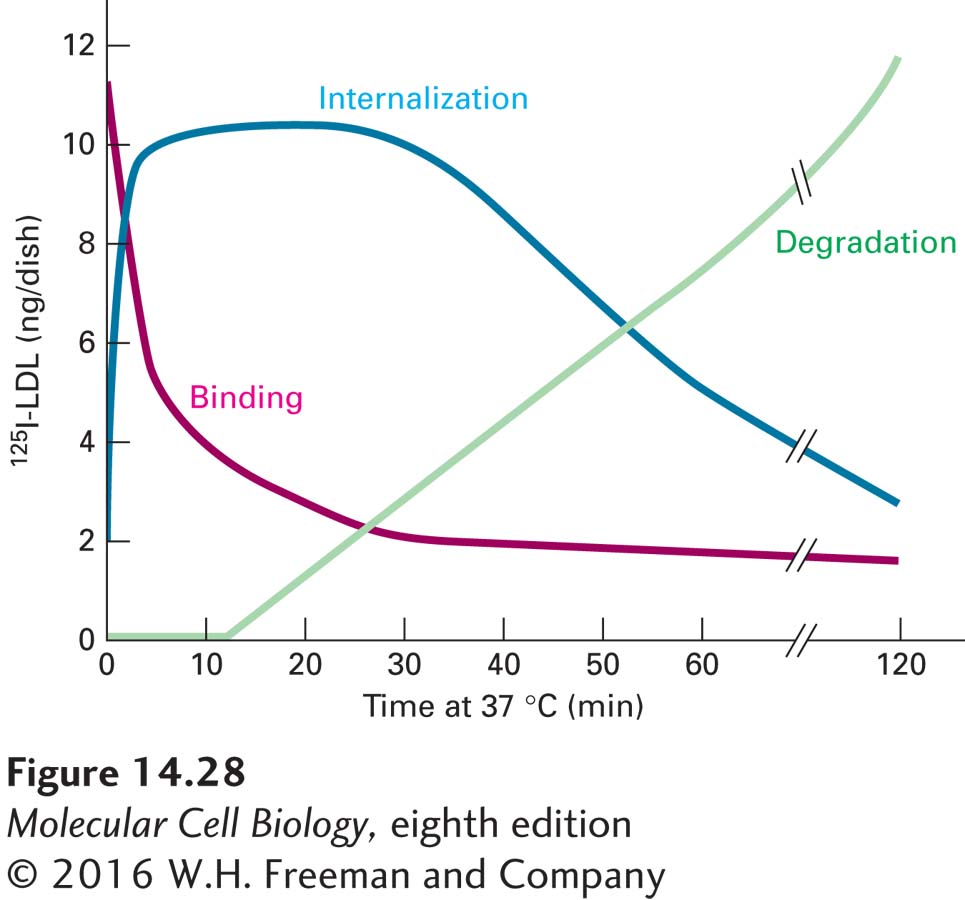
Two general experimental approaches have been used to study how LDL particles enter cells. The first method makes use of LDL that has been labeled by the covalent attachment of radioactive 125I to the side chains of tyrosine residues in apoB-100 on the surfaces of the LDL particles. After cultured cells are incubated for several hours with the labeled LDL, it is possible to determine how much LDL is bound to the surfaces of cells, how much is internalized, and how much of the apoB-100 component of the LDL is degraded by enzymatic hydrolysis to individual amino acids. The degradation of apoB-100 can be detected by the release of 125I-tyrosine into the culture medium. Figure 14-28 shows the time course of these events in receptor-mediated cellular LDL processing, determined by pulse-chase experiments with a fixed concentration of 125I-labeled LDL. These experiments clearly demonstrate the order of events: surface binding of LDL → internalization → degradation. The second approach involves tagging LDL particles with an electron-dense label that can be detected by electron microscopy. Such studies can reveal the details of how LDL particles first bind to the surface of cells at clathrin-coated endocytic pits, then remain associated with the coated pits as they invaginate and bud off to form coated vesicles and are finally transported to endosomes (see Figure 14-26).
Page 660
EXPERIMENTAL FIGURE 14-28Pulse-chase experiment demonstrates precursor-product relations in cellular uptake of LDL. Cultured normal human skin fibroblasts were incubated in a medium containing 125I-LDL for 2 hours at 4 °C (the pulse). After excess 125I-LDL not bound to the cells was washed away, the cells were incubated at 37 °C for the indicated amounts of time in the absence of external LDL (the chase). The amounts of surface-bound, internalized, and degraded (hydrolyzed) 125I-LDL were measured. Binding, but not internalization or hydrolysis, of LDL apoB-100 occurs during the 4 °C pulse. The data show the very rapid disappearance of bound 125I-LDL from the surface as it is internalized after the cells have been warmed to allow membrane movements. After a lag period of 15–20 minutes, lysosomal degradation of the internalized 125I-LDL commences. See M. S. Brown and J. L. Goldstein, 1976, Cell9:663.