All G Protein–Coupled Receptors Share the Same Basic Structure
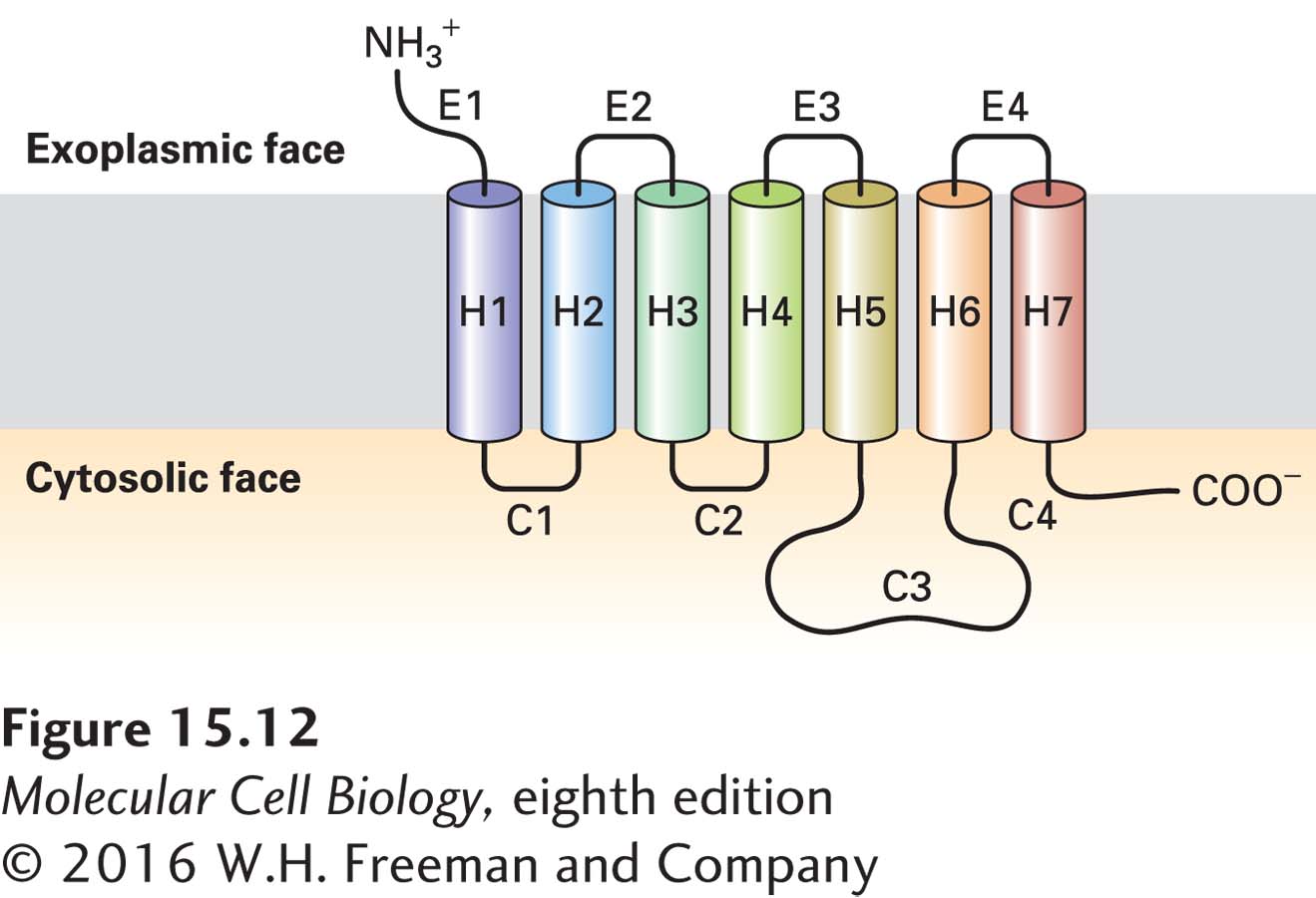
All G protein–coupled receptors have the same orientation in the membrane and contain seven transmembrane α-helical regions (H1–H7), four extracellular segments, and four cytosolic segments (Figure 15-12). Invariably the N-terminus is on the exoplasmic face and the C-terminus is on the cytosolic face of the plasma membrane. The human genome encodes some 800 functional GPCRs, which are divided into several subfamilies; members of these subfamilies are especially similar in amino acid sequence and structure.
FIGURE 15-12General structure of G protein–coupled receptors. All receptors of this type have the same orientation in the membrane and contain seven transmembrane α-helical regions (H1–H7), four extracellular segments (E1–E4), and four cytosolic segments (C1–C4).
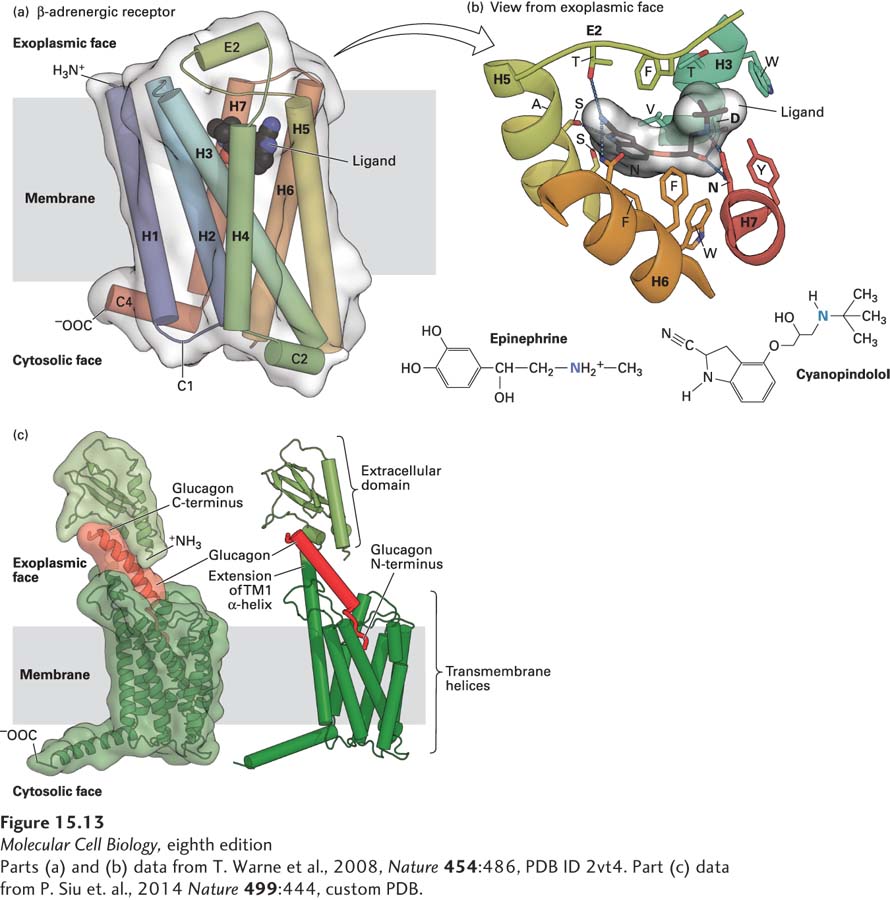
G protein–coupled receptors are stably anchored in the hydrophobic core of the plasma membrane by the many hydrophobic amino acids on the outer surfaces of the seven membrane-spanning segments. One group of G protein–coupled receptors whose structure is known in molecular detail is the β-adrenergic receptors, which bind hormones such as epinephrine and norepinephrine (Figure 15-13a). In these and many other receptors, segments of several membrane-embedded α helices and extracellular loops form the ligand-binding site, which is open to the exoplasmic surface. The antagonist cyanopindolol, shown in Figure 15-13b, binds with a much higher affinity to the β-adrenergic receptor than do most agonists, and the resulting receptor-ligand complex has been crystallized and its structure determined. Side chains of 15 amino acids located in four transmembrane α helices and extracellular loop E2 make noncovalent contacts with the ligand.
[Parts (a) and (b) data from T. Warne et al., 2008, Nature454:486, PDB ID 2vt4. Part (c) data from P. Siu et al., 2014 Nature499:444, custom PDB.]
FIGURE 15-13Binding of ligands to GPCRs. (a) Structure of the turkey β1-adrenergic receptor bound to its antagonist cyanopindolol. This side view shows the approximate location of the membrane phospholipid bilayer. The ribbon representation of the receptor structure is in rainbow coloration (N-terminus, blue; C-terminus, red), with cyanopindolol as a gray space-filling molecule. The extracellular loop 2 (E2) and cytoplasmic loops 1, 2, and 4 (C1, C2, C4) 1 and 2 (C1, C2) are labeled. (b) The hormone-binding pocket formed by residues in several transmembrane segments. View from external face showing a close-up of the ligand-binding pocket that is formed by amino acids in helices 3, 5, 6, and 7, as well as extracellular loop 2, located between helices 4 and 5. Cyanopindolol atoms are colored gray (carbon), blue (nitrogen), and red (oxygen). The ligand-binding pocket comprises 15 side chains from amino acid residues in four transmembrane α helices and extracellular loop 2. As examples of specific binding interactions, the positively charged N atom in the amino group found both in cyanopindolol and in epinephrine forms an ionic bond with the carboxylate side chain of aspartate 121 (D) in helix 3 and the carboxylate of asparagine 329 (N) in helix 7. (c) Model for glucagon binding to the glucagon receptor. The seven transmembrane α helices of the glucagon receptor and the exoplasmic extension of transmembrane helix 1 are colored dark green and the N-terminal exoplasmic domain light green. The C-terminus of the 29-amino-acid peptide glucagon (red) is bound to the receptor N-terminal domain, and the glucagon N-terminus is thought to insert into a binding pocket that is in the center of the seven transmembrane α helices.
[Parts (a) and (b) data from T. Warne et al., 2008, Nature454:486, PDB ID 2vt4. Part (c) data from P. Siu et al., 2014 Nature499:444, custom PDB.]
Page 687
Glucagon is a 29-amino-acid peptide hormone secreted by the α cells of the pancreatic islets; as we will see later in this chapter, it acts on the liver to trigger glycogen breakdown and secretion of glucose into the bloodstream. The glucagon receptor is in a different GPCR subfamily from the β-adrenergic receptors; while it has the standard seven transmembrane α helices, it also has a large exoplasmic domain that is connected to an extension of the first transmembrane α helix. This exoplasmic domain binds tightly to the C-terminus of glucagon, positioning the N-terminus to bind to the pocket formed, as in β-adrenergic and other GPCRs, by residues from several transmembrane helices (Figure 15-13c). The amino acids that form the interior of different GPCRs are diverse, allowing different receptors to bind very different small molecules, whether they are hydrophilic, such as epinephrine and glucagon, or hydrophobic, such as many odorants. But in all cases, binding of the ligand causes a conformational change in the receptor that enables it to activate the GTP-binding Gα subunit of a heterotrimeric G protein.