The human retina contains two types of photoreceptor cells, rods and cones, which are the primary recipients of visual stimulation. Cones are involved in color vision, while rods are stimulated by weak light such as moonlight over a range of wavelengths. The photoreceptor cells signal to (synapse with) layer upon layer of interneurons that are innervated by different combinations of photoreceptor cells. All these signals are processed and relayed through the visual thalamus to the part of the brain called the visual cortex, where they are interpreted.
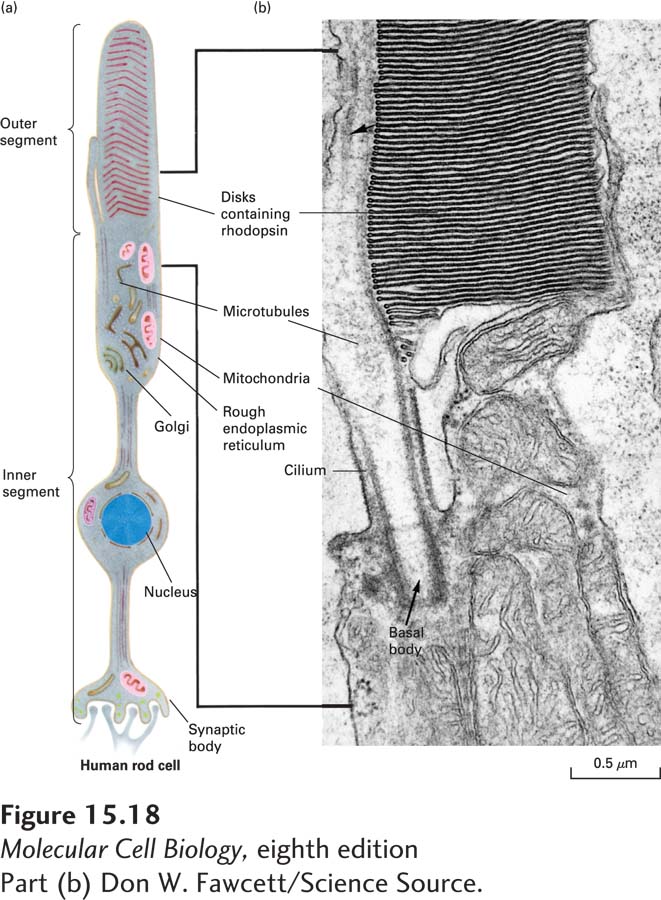
Rod cells sense light with the aid of a light-sensitive GPCR known as rhodopsin. Rhodopsin consists of the protein opsin, which has the usual seven–transmembrane segment GPCR structure, covalently linked to a light-absorbing pigment called retinal. Rhodopsin, found only in rod cells, is localized to the thousand or so flattened membrane disks that make up the outer segment of each of these rod-shaped cells (Figure 15-18). A human rod cell contains about 4 × 107 molecules of rhodopsin. The heterotrimeric G protein coupled to rhodopsin, called transducin (Gt), contains a Gα unit referred to as Gαt; like rhodopsin, Gαt is found only in rod cells.
[Part (b) Don W. Fawcett/Science Source.]
FIGURE 15-18Human rod cell. (a) Schematic diagram of an entire rod cell. At the synaptic body, the rod cell forms synapses with one or more interneurons. Rhodopsin, a light-sensitive G protein–coupled receptor, is located in the flattened membrane disks of the cell’s outer segment. (b) Electron micrograph of the region of the rod cell indicated by the bracket in (a). This region includes the junction of the inner and outer segments.
[Part (b) Don W. Fawcett/Science Source.]
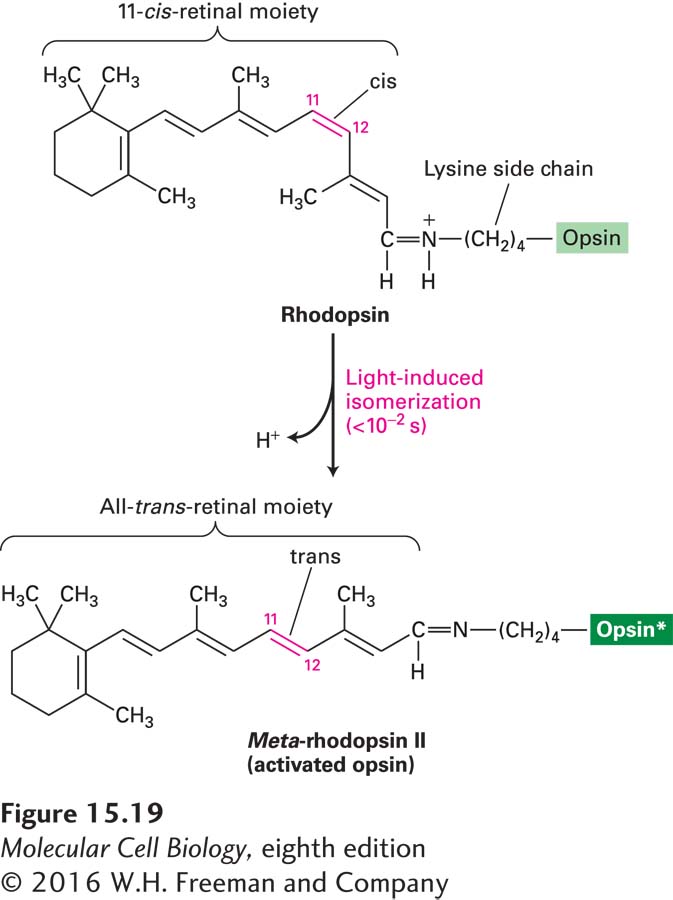
Rhodopsin (R) differs from other GPCRs in that binding of a ligand is not what activates the receptor. Rather, absorption of a photon of light by the bound retinal is the activating signal. On absorption of a photon, the retinal moiety of rhodopsin is immediately converted from the cis isomeric form (known as 11-cis-retinal) to the all-trans isomeric form, causing a conformational change in the opsin protein (Figure 15-19). This change is equivalent to the activating conformational change that occurs upon ligand binding by other G protein–coupled receptors; this conformational change allows rhodopsin to bind the Gαt subunit of its coupled G protein, transducin, triggering exchange of GTP for GDP on its Gαt subunit. At the same time, transducin also dissociates into its Gαt and Gβγ subunits. The activated rhodopsin that is formed, termed R*, is unstable, as the covalent linkage to retinal is spontaneously cleaved. Since retinal-free opsin cannot bind transducin, initiation of visual signaling is terminated at this point. In the dark, free all-trans-retinal is converted back to 11-cis-retinal in a series of steps involving enzymes from both rod cells and cells in the adjacent retinal pigment epithelium. The 11-cis-retinal that is generated is then transferred to the rod cells, where it rebinds opsin, forming rhodopsin and completing the rhodopsin visual cycle.
Page 695
FIGURE 15-19Vision depends on the light-triggered isomerization of the retinal moiety of rhodopsin. Rhodopsin consists of the light-absorbing pigment 11-cis-retinal covalently attached to the amino group of lysine residue 296 in the opsin protein. Absorption of light causes rapid photoisomerization of the bound cis-retinal to the all-trans isomer. This change triggers a conformational change in rhodopsin, forming the unstable intermediate meta-rhodopsin II, or activated rhodopsin (see Figure 15-21), which activates Gt proteins. Within seconds, all-trans-retinal dissociates from opsin and is converted by a series of enzymes in the rod cell and pigmented epithelium back to the cis isomer, which then rebinds to another opsin molecule. See J. Nathans, 1992, Biochemistry31:4923.