Rapid Termination of the Rhodopsin Signal Transduction Pathway Is Essential for the Temporal Resolution of Vision
As in all G protein–coupled signaling pathways, timely termination of the rhodopsin signaling pathway requires that all the activated intermediates be inactivated rapidly, restoring the system to its basal state, ready for signaling again. Thus the three protein intermediates, activated rhodopsin (R*), Gαt·GTP, and activated PDE, must all be inactivated, and the concentration of cytoplasmic cGMP must be restored to its dark level by guanylyl cyclase, the enzyme that catalyzes production of cGMP from GMP. The entire process of rhodopsin activation and inactivation in a mammalian rod cell as it responds to a single photon of light takes only about 50 milliseconds; this enables the eye to respond very quickly to changing light conditions. Several mechanisms act together to make possible this very rapid response.
A GTPase Activating Protein (GAP) That Inactivates Gαt·GTP First, the complex composed of the inhibitory γ subunit of PDE and Gαt·GTP recruits two additional proteins, RGS9 and Gβ5, that together act as a GAP to enhance the rate of hydrolysis of bound GTP to GDP (step 7, Figure 15-20). Hydrolysis of GTP, in turn, causes release of the PDE γ subunit, which rejoins the PDE α and β subunits, terminating PDE activation (step 8). Experiments with mice in which the gene encoding RGS9 had been knocked out showed that this protein is essential for normal inactivation of the signaling cascade in vivo. In individual mouse rod cells, the time required for recovery from a single flash of light increased from 0.2 seconds in the normal mouse to about 9 seconds in the RGS9-deficient mouse. This represents a 45-fold increase, attesting to the importance of RGS9 as a component of the Gαt·GTP-GAP complex.
Ca2+-Sensing Proteins That Activate Guanylate Cyclase Second, light-triggered closing of the cGMP-gated cation channels causes a drop in the cytosolic Ca2+ concentration inside the rod cell. This happens because Ca2+ is continually being transported out of the cells by a type of antiporter protein (the Na+/Ca2+-K+ exchanger) that is not affected by cGMP levels. The drop in the intracellular Ca2+ concentration is sensed by a class of Ca2+-binding proteins known as guanylate cyclase–activating proteins, which bind to guanylate cyclase and stimulate its activity, thereby elevating the level of cGMP and causing the cGMP-gated ion channels to reopen. This is another example of negative feedback, in which a downstream second message—in this case, low cytosolic Ca2+—acts to inhibit further signaling and thus prevent an overreaction by the cell.
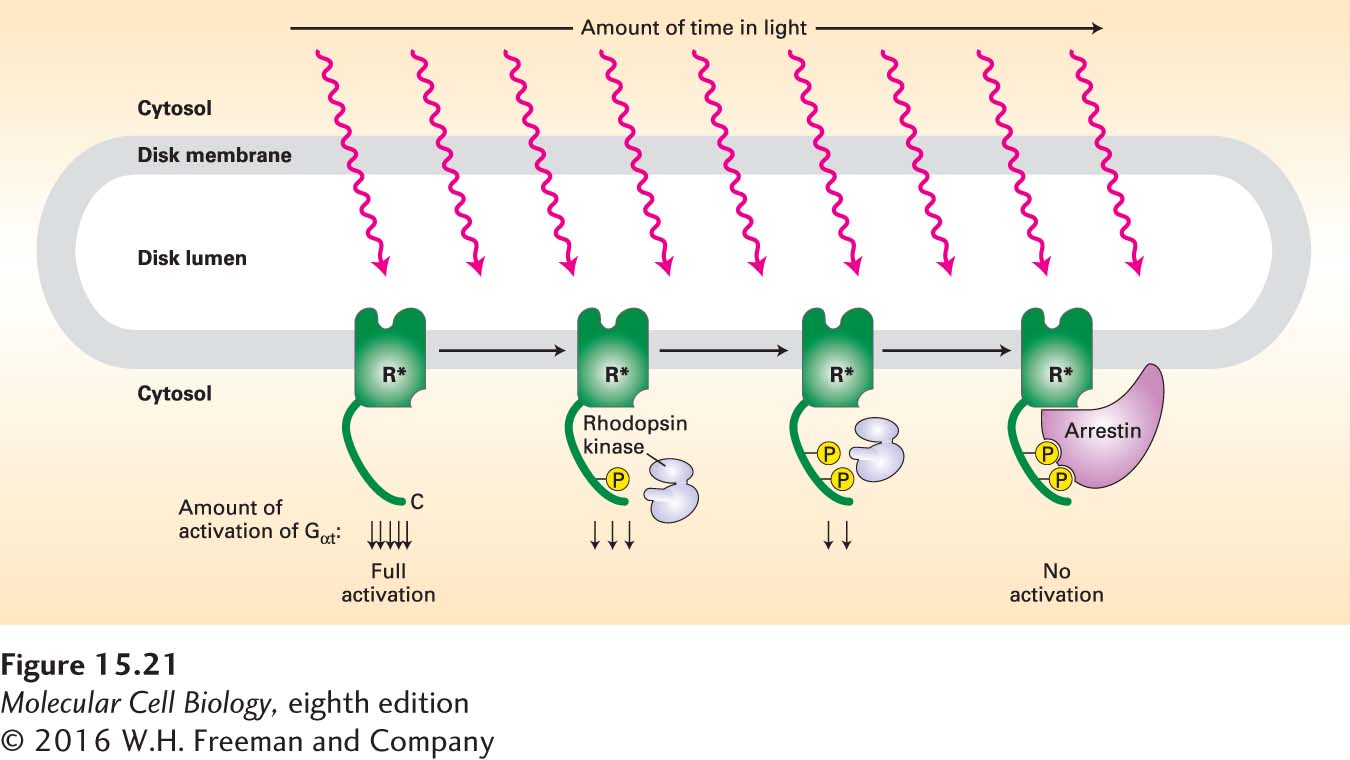
Rhodopsin Phosphorylation and Binding of Arrestin Third, a major process that suppresses and helps to terminate the visual response involves phosphorylation of rhodopsin when it is in its activated (R*), but not its inactivated, or dark (R), form. Rhodopsin kinase (Figure 15-21), a member of a class of GPCR kinases, is the enzyme that catalyzes this phosphorylation reaction. Each opsin molecule has two principal serine phosphorylation sites on its cytosol-facing C-terminal C4 segment; the more sites that are phosphorylated by rhodopsin kinase, the less able R* is to activate Gαt and so induce the closing of cGMP-gated cation channels.
FIGURE 15-21Inhibition of rhodopsin signaling by rhodopsin kinase. Light-activated rhodopsin (R*), but not dark-adapted rhodopsin, is a substrate for rhodopsin kinase. The extent of rhodopsin phosphorylation is proportional to the amount of time each rhodopsin molecule spends in the light-activated form; the greater the extent of the phosphorylation, the more the ability of R* to activate transducin is reduced. Arrestin binds to the completely phosphorylated opsin, forming a complex that cannot activate transducin at all. See A. Mendez et al., 2000, Neuron28:153, and V. Arshavsky, 2002, Trends Neurosci.25:124.
Page 698
The protein arrestin, which binds tightly to rhodopsin only when two or three of the serines are phosphorylated, dramatically speeds up the inactivation process. Arrestin bound to the phosphorylated R* completely prevents interaction with Gαt, blocking formation of the active Gαt·GTP complex and stopping further activation of PDE. The entire process of rhodopsin phosphorylation and inactivation by arrestin is completed very quickly, within 50 milliseconds. Meanwhile, the phosphates linked to rhodopsin are continuously being removed by a specific rhodopsin phosphatase; this removal causes the dissociation of arrestin and the restoration of rhodopsin to its original, light-sensitive state.