Signaling Molecules Can Act Locally or at a Distance
As noted above, cells respond to many different types of signals—
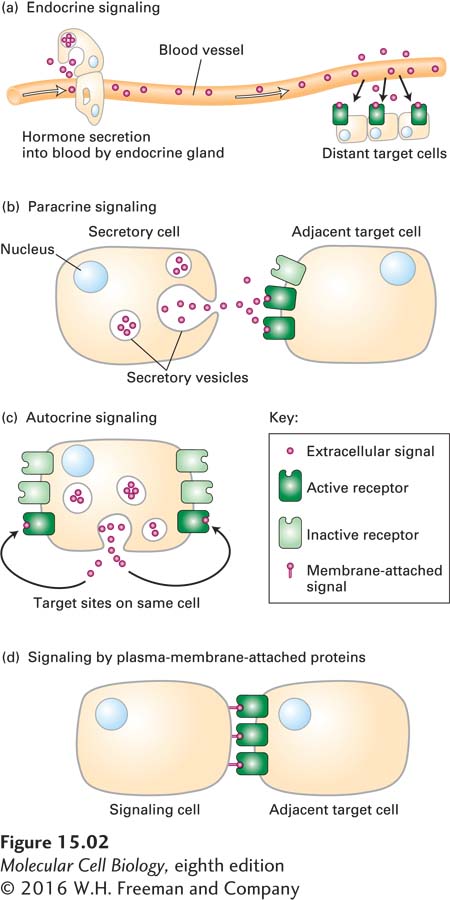
Page 676
In endocrine signaling, the signaling molecules are synthesized and secreted by signaling cells (for example, those found in endocrine glands), transported through the circulatory system of the organism, and finally act on target cells distant from their site of synthesis. The term hormone generally refers to signaling molecules that mediate endocrine signaling. Insulin secreted by the pancreas and epinephrine secreted by the adrenal glands are examples of hormones that travel through the blood and thus mediate endocrine signaling.
In paracrine signaling, the signaling molecules released by a cell affect only those target cells in close proximity. A neuron releasing a neurotransmitter (e.g., acetylcholine) that acts on an adjacent neuron or on a muscle cell (inducing or inhibiting muscle contraction) is an example of paracrine signaling. In addition to neurotransmitters, many of the protein growth factors that regulate development in multicellular organisms act at short range. Some of these proteins bind tightly to components of the extracellular matrix and are unable to signal to adjacent cells; subsequent degradation of these matrix components, triggered by injury or infection, releases these growth factors and enables them to signal. Many of the developmentally important signaling proteins that we discuss in Chapter 16 diffuse away from the signaling cell, forming a concentration gradient and inducing different responses in adjacent cells depending on the concentration of the signaling protein.
In autocrine signaling, cells respond to substances that they themselves release. Some growth factors act in this fashion, and cultured cells often secrete growth factors that stimulate their own growth and proliferation. This type of signaling is particularly characteristic of tumor cells, many of which overproduce and release growth factors that stimulate inappropriate, unregulated self-
Many integral membrane proteins located on the cell surface play important roles as signals (Figure 15-2d). In some cases, such membrane-
Some signaling molecules can act at both short and long ranges. For example, epinephrine (also known as adrenaline) functions as a hormone (endocrine signaling) as part of the “fight or flight” response to a sudden danger in the environment, and also as a neurotransmitter (paracrine signaling). Another example, discussed in Chapter 16, is epidermal growth factor (EGF), which is synthesized as an integral plasma-