The Infancy of Signal Transduction Studies: GTP Stimulation of cAMP Synthesis
M. Rodbell et al., 1971, J. Biol. Chem.246:1877
In the late 1960s, the study of hormone action blossomed following the discovery that cyclic adenosine monophosphate (cAMP) functions as a second messenger, coupling the hormone-mediated activation of a receptor to a cellular response. In setting up an experimental system to investigate the hormone-induced synthesis of cAMP, Martin Rodbell discovered an important new player in intracellular signaling: guanosine triphosphate (GTP).
Background
The discovery of GTP’s role in regulating signal transduction began with studies on how glucagon and other hormones send a signal across the plasma membrane that eventually evokes a cellular response. At the outset of Rodbell’s studies, it was known that binding of glucagon to specific receptor proteins embedded in the membrane stimulates production of cAMP. It was also known that the formation of cAMP from ATP is catalyzed by a membrane-bound enzyme called adenylyl cyclase. It had been proposed that the action of glucagon, and other cAMP-stimulating hormones, relies on additional molecular components that couple receptor activation to the production of cAMP. However, in studies with isolated fat-cell plasma membranes known as “ghosts,” Rodbell and his coworkers were unable to provide any further insight into how glucagon binding leads to an increase in production of cAMP. Rodbell then began a series of studies with a newly developed cell-free system, purified rat liver plasma membranes, which retained both membrane-bound and membrane-associated proteins. These experiments eventually led to the finding that GTP is required for the glucagon-induced stimulation of adenylyl cyclase.
The Experiment
One of Rodbell’s first goals was to characterize the binding of glucagon to the glucagon receptor in the cell-free rat liver membrane system. First, purified rat liver membranes were incubated with glucagon labeled with the radioactive isotope of iodine (125I). The membranes were then separated from the unbound [125I] glucagon by centrifugation. Once it was established that labeled glucagon would indeed bind to the membranes, the study went on to determine if this binding led directly to activation of adenylyl cyclase and production of cAMP in the membranes.
The production of cAMP in the cell-free system required the addition of three components: ATP; the substrate for adenylyl cyclase, Mg2+; and an ATP-regenerating system consisting of creatine kinase and phosphocreatine. Surprisingly, when the glucagon-binding experiment was repeated in the presence of these additional factors, Rodbell observed a 50 percent decrease in glucagon binding. Full binding could be restored only when ATP was omitted from the reaction. This observation inspired an investigation of the effect of nucleoside triphosphates on the binding of glucagon to its receptor. It was shown that relatively high (i.e., millimolar) concentrations not only of ATP, but also of uridine triphosphate (UTP) and cytidine triphosphate (CTP), reduced the binding of labeled glucagon. In contrast, the reduction of glucagon binding in the presence of GTP occurred at far lower (micromolar) concentrations. Moreover, low concentrations of GTP were found to stimulate the dissociation of bound glucagon from the receptor. Taken together, these studies suggested that GTP alters the glucagon receptor in a manner that lowers its affinity for glucagon. This decreased affinity both affects the ability of glucagon to bind to the receptor and encourages the dissociation of bound glucagon.
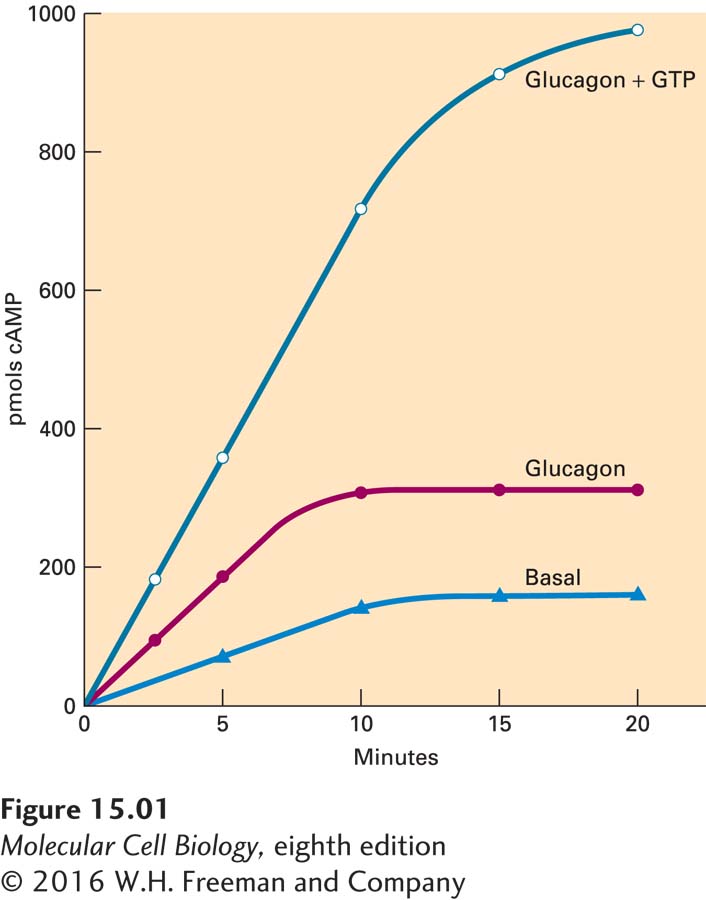
The observation that GTP was involved in the action of glucagon led to a second key question: Can GTP also exert an effect on adenylyl cyclase? Addressing this question experimentally required the addition of both ATP, as a substrate for adenylyl cyclase, and GTP, as the factor being examined, to the purified rat liver membranes. However, the previous study had shown that the concentration of ATP required as a substrate for adenylyl cyclase could affect glucagon binding. Might it also activate adenylyl cyclase? The concentration of ATP used in the experiment could not be reduced because ATP was readily hydrolyzed by ATPases present in the rat liver membrane. To get around this dilemma, Rodbell replaced ATP with an AMP analog, 5′-adenyl-imidodiphosphate (AMP-PNP), which can be converted to cAMP by adenylyl cyclase but is resistant to hydrolysis by membrane ATPases. The critical experiment could now be performed. Purified rat liver membranes were treated with glucagon both in the presence and in the absence of GTP, and the production of cAMP from AMP-PNP was measured. The addition of GTP clearly stimulated the production of cAMP when compared with the addition of glucagon alone (Figure 1), indicating that GTP not only affects the binding of glucagon to its receptor, but also stimulates the activation of adenylyl cyclase.
FIGURE 1Effect of GTP on glucagon-stimulated cAMP production from AMP-PNP by purified rat liver membranes.
Discussion
Two key factors led Rodbell and his colleagues to detect the role of GTP in signal transduction, whereas previous studies had failed to do so. First, by switching from fat-cell ghosts to the rat liver membrane system, the Rodbell team avoided contamination of their cell-free system with GTP, a problem associated with the procedure for isolating ghosts. Such contamination would have masked the effects of GTP on glucagon binding and activation of adenylyl cyclase. Second, when ATP was first shown to influence glucagon binding, Rodbell did not simply accept the plausible explanation that ATP, the substrate for adenylyl cyclase, also affects binding of glucagon. Instead, he chose to test the effects of the other common nucleoside triphosphates on binding. Rodbell later noted that he knew commercial preparations of ATP are often contaminated with low concentrations of other nucleoside triphosphates. The possibility of contamination suggested to him that small concentrations of GTP might exert large effects on glucagon binding and the stimulation of adenylyl cyclase.
This critical series of experiments stimulated a large number of studies on the role of GTP in hormone action, eventually leading to the discovery of G proteins, the GTP-binding proteins that couple certain receptors to adenylyl cyclase. Subsequently, an enormous family of receptors that require G proteins to transduce their signals were identified in eukaryotes from yeast to humans. These G protein–coupled receptors are involved in the action of many hormones as well as in a number of other biological activities, including neurotransmission and the immune response. It is now known that binding of ligands to their cognate G protein–coupled receptors stimulates the associated G proteins to bind GTP. This binding causes transduction of a signal that stimulates adenylyl cyclase to produce cAMP as well as desensitization of the receptor, which then releases its ligand. Both of these effects were observed in Rodbell’s experiments on glucagon action. For these seminal observations, Rodbell was awarded the Nobel Prize in Physiology or Medicine in 1994.