Signals Pass from Activated Ras to a Cascade of Protein Kinases Ending with MAP Kinase
Biochemical and genetic studies in yeast, C. elegans, Drosophila, and mammals have revealed that downstream of Ras is a highly conserved cascade of three protein kinases, culminating in MAP kinase. Although activation of the kinase cascade does not yield the same biological results in all cells, a common set of sequentially acting kinases defines the Ras/MAP kinase pathway, as outlined in Figure 16-24: Ras → Raf → MEK → MAP kinase.
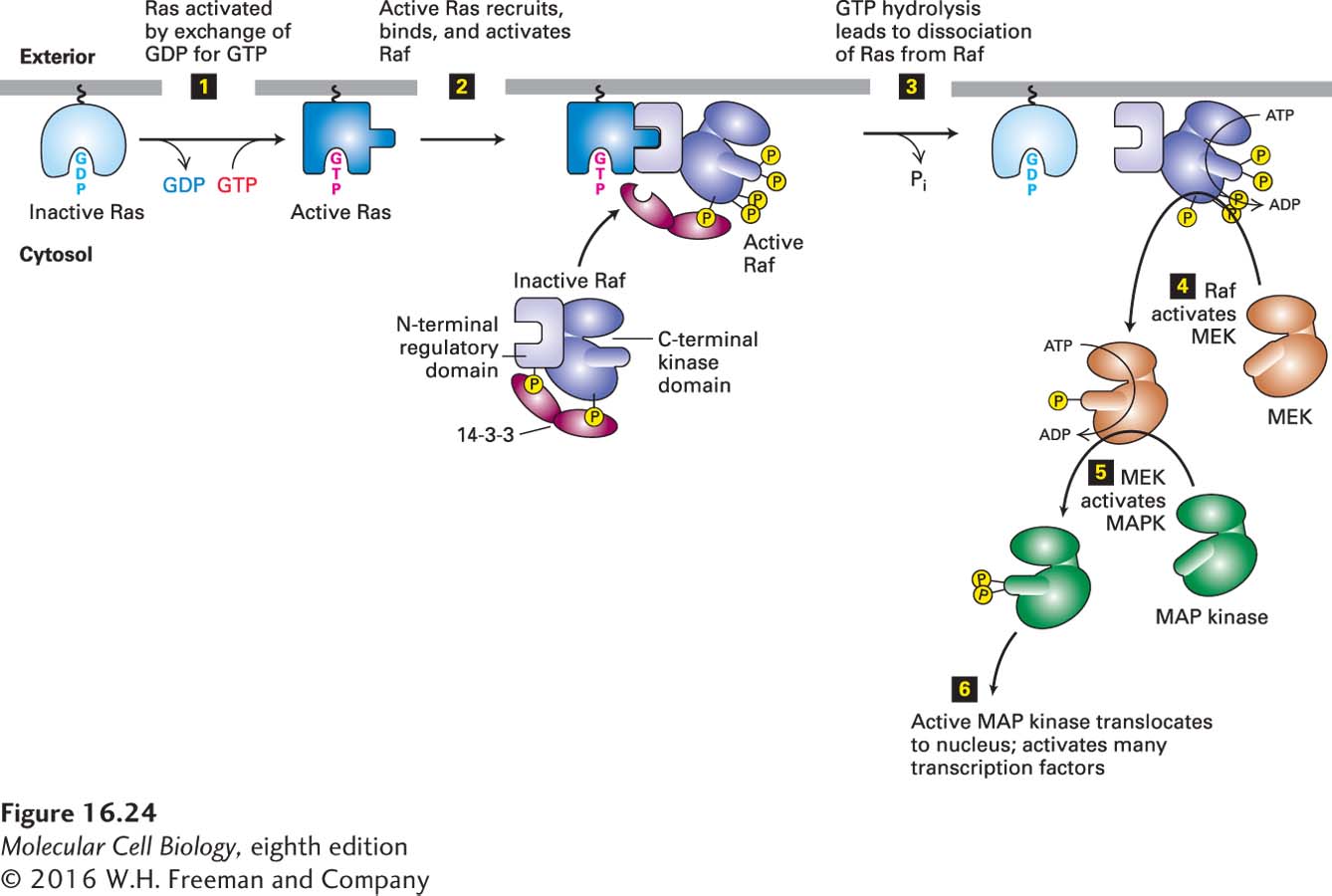
FIGURE 16-24Ras/MAP kinase pathway. In unstimulated cells, most Ras is tethered to the cytosolic surface of the plasma membrane in the inactive form with bound GDP. Binding of a ligand to its RTK or cytokine receptor leads to formation of the active Ras·GTP complex (step 1; see also Figure 16-21). Activated Ras triggers the downstream kinase cascade depicted in steps 2–6, culminating in activation of MAP kinase (MAPK). In unstimulated cells, binding of a dimer of the 14-3-3 protein to Raf stabilizes it in an inactive conformation. Each 14-3-3 monomer binds to a phosphoserine residue in Raf, one to phosphoserine-259 in the N-terminal domain and the other to phosphoserine-621 in the kinase domain; binding to 14-3-3 maintains the kinase in a closed, inactive state. Interaction of the Raf N-terminal regulatory domain with Ras·GTP results in dephosphorylation of one of the serines that bind Raf to 14-3-3, phosphorylation of other residues, loss of 14-3-3 binding, and activation of Raf kinase activity. After inactive Ras·GDP dissociates from Raf, it can presumably be reactivated by signals from activated receptors, thereby recruiting additional Raf molecules to the membrane. See E. Kerkhoff and U. Rapp, 2001, Adv. Enzyme Regul.41:261; J. Avruch et al., 2001, Recent Prog. Horm. Res.56:127; and D. Matallanas et al., 2011, Genes Cancer2:232.
Ras is activated by the exchange of GDP for GTP (step 1). Active Ras·GTP binds to the N-terminal regulatory domain of Raf, a serine/threonine (not tyrosine) kinase, thereby activating its kinase activity (step 2). In unstimulated cells, Raf is phosphorylated and bound in an inactive state by the protein 14-3-3, which binds phosphoserine residues in a number of important signaling proteins. Hydrolysis of Ras · GTP to Ras · GDP releases active Raf from its complex with 14-3-3 (step 3), and the now active Raf subsequently phosphorylates and thereby activates MEK (step 4). MEK, a dual-specificity protein kinase, phosphorylates its target proteins on both tyrosine and serine/threonine residues. Active MEK mainly phosphorylates and activates MAP kinase, another serine/threonine kinase also known as ERK (step 5). In different cells, MAP kinase phosphorylates many different proteins, including nuclear transcription factors that mediate cellular responses (step 6).
Page 743
Page 744
Several types of experiments have demonstrated that Raf, MEK, and MAP kinase lie downstream from Ras and have revealed the sequential order of these proteins in the pathway. For example, cultured mammalian cells that express a mutant, nonfunctional Raf protein cannot be stimulated to proliferate uncontrollably by a constitutively active RasD protein. This finding established a link between the Raf and Ras proteins and showed that Raf lies downstream of Ras in the signaling pathway. In vitro binding studies further showed that the purified Ras·GTP protein binds directly to the N-terminal regulatory domain of Raf and activates its catalytic activity.
That MAP kinase is activated in response to Ras activation was demonstrated in quiescent cultured cells expressing a constitutively active RasD protein. In these cells, activated MAP kinase is generated in the absence of stimulation by growth-promoting hormones. More importantly, R7 photoreceptors develop normally in the developing eyes of Drosophila mutants that lack a functional Ras or Raf protein but express a constitutively active MAP kinase. This finding indicates that activation of MAP kinase is sufficient to transmit a proliferation or differentiation signal normally initiated by ligand binding to a receptor tyrosine kinase such as Sevenless (see Figure 16-20). Biochemical studies showed, however, that Raf cannot directly phosphorylate MAP kinase or otherwise activate its activity.
The final link in the kinase cascade activated by Ras·GTP emerged from studies in which scientists fractionated extracts of cultured cells to search for a kinase activity that could phosphorylate MAP kinase and that was present only in cells stimulated with growth factors, not in unstimulated cells. This work led to the identification of MEK, a kinase that specifically phosphorylates one threonine and one tyrosine residue on the activation loop of MAP kinase, thereby activating its catalytic activity. (The acronym MEK comes from MAP and ERK kinase.) Later studies showed that MEK binds to the C-terminal catalytic domain of Raf and is phosphorylated by the Raf serine/threonine kinase; this phosphorylation activates the catalytic activity of MEK.
Hence activation of Ras induces a kinase cascade that includes Raf, MEK, and MAP kinase: activated RTK → Ras → Raf → MEK → MAP kinase. Although we will not emphasize them here, the complexity of this pathway is increased by the multiple isoforms of each of its components. In humans, there are three RAS, three Raf, two MEK, and two ERK proteins, and each of these has overlapping but also nonredundant functions.
Activating mutations in the Raf gene occur in about 50 percent of melanomas, skin cancers that are often caused by exposure to the ultraviolet radiation in sunlight. Among these melanomas, one particular mutation, a glutamic acid substitution for the valine at position 600, occurs in over 90 percent. This mutant Raf stimulates MEK-ERK signaling in cells in the absence of growth factors, and mutant Raf transgenes induce melanoma in mice. Very potent and selective inhibitors of Raf have recently entered the clinic and are producing excellent responses in patients with melanomas caused by this mutant Raf protein.