G Protein–Coupled Receptors Transmit Signals to MAP Kinase in Yeast Mating Pathways
Although MAP kinase is often activated in multicellular animals by RTKs or cytokine receptors, signaling from other receptors can activate MAP kinase in other eukaryotic cells (see Figure 15-32). To illustrate, we consider the mating pathway in the yeast S. cerevisiae, a well-studied example of a MAP kinase cascade linked to G protein–coupled receptors (GPCRs), in this case for two secreted peptide pheromones, the a and α factors.
Haploid yeast cells are either of the a or α mating type. They secrete protein signals known as pheromones, which induce mating between haploid yeast cells of the opposite mating types. An a haploid cell secretes the a mating factor and has GPCRs for the α factor on its cell surface; an α cell secretes the α factor and has GPCRs for the a factor (see Figure 1-23). Thus each type of cell recognizes the mating factor produced by the opposite type. Activation of the MAP kinase pathway by either the a or the α GPCRs induces transcription of genes that inhibit progression of the cell cycle and others that enable cells of opposite mating type to fuse together and ultimately form a diploid cell.
Ligand binding to either of the two yeast pheromone GPCRs triggers the exchange of GTP for GDP on the Gα subunit and dissociation of Gα·GTP from the Gβγ complex. This activation process is identical to that for the GPCRs discussed in the previous chapter (see Figure 15-14). In many mammalian GPCR-initiated pathways, the active Gα transduces the signal. In contrast, the dissociated Gβγ complex mediates all the physiological responses induced by activation of the yeast pheromone GPCRs (Figure 16-27a). As evidence, in yeast cells that lack Gα, the Gβγ subunit is always free. Such cells can mate in the absence of mating factors; that is, the mating response is constitutively on. However, in cells defective for the Gβ or Gγ subunit, the mating pathway cannot be induced at all. If dissociated Gα were the transducer, the pathway would be expected to be constitutively active in these mutant cells.
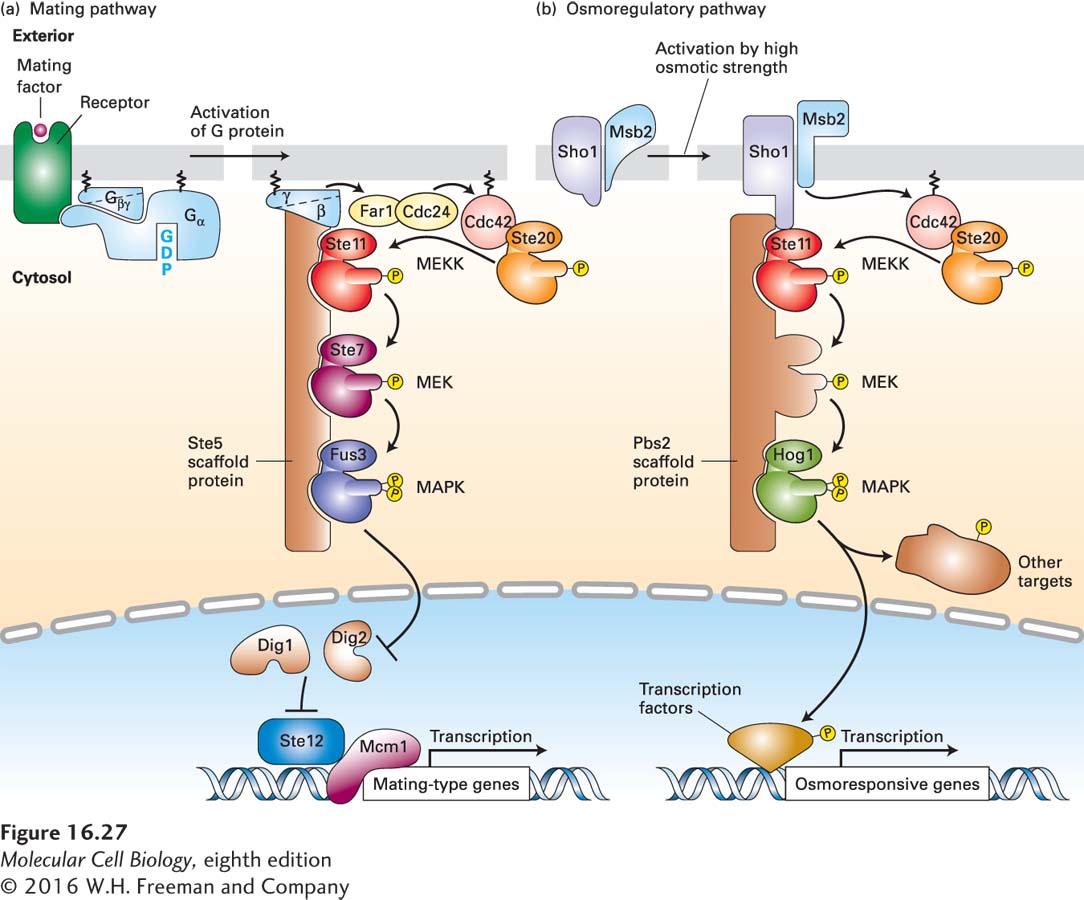
FIGURE 16-27Scaffold proteins separate yeast MAP kinase cascades in the mating and osmoregulatory pathways. In yeast, different receptors activate different MAP kinase pathways, two of which are outlined here. The two MEKs depicted, like all MEKs, are dual-specificity threonine/tyrosine kinases; all of the other kinases are serine/threonine kinases. (a) Mating pathway: The receptors for yeast α and a mating factors are coupled to the same trimeric G protein. Following ligand binding and dissociation of the G protein subunits, the membrane-tethered Gβγ subunit binds the Ste5 scaffold to the plasma membrane. Gβγ also activates Cdc24, a GEF for the Ras-like protein Cdc42; the active, GTP-bound Cdc42, in turn, binds to and activates the Ste20 kinase. Ste20 then phosphorylates and activates Ste11, which is analogous to Raf and other mammalian MEK kinase (MEKK) proteins. Ste20 thus serves as a MAPKKK kinase. Ste11 initiates a kinase cascade in which the final component, Fus3, is functionally equivalent to MAP kinase (MAPK) in higher eukaryotes. Like other MAP kinases, activated Fus3 then translocates into the nucleus. There it phosphorylates two proteins, Dig1 and Dig2, relieving their inhibition of the Ste12 transcription factor, allowing it to bind to DNA and initiate transcription of genes that inhibit progression of the cell cycle and others that enable cells of opposite mating type to fuse together and ultimately form a diploid cell. (b) Osmoregulatory pathway: Two plasma membrane proteins, Sho1 and Msb1, are activated in an unknown manner by exposure of yeast cells to media of high osmotic strength. Activated Sho1 recruits the Pbs2 scaffold protein, which contains a MEK domain, to the plasma membrane. At the plasma membrane, the Sho1-Msb1 complex activates Cdc42, which in turn activates the resident Ste20 kinase, as in the mating pathway. Ste20, in turn, phosphorylates and activates Ste11, initiating a kinase cascade that activates Hog1, a MAP kinase. In the cytosol, Hog1 phosphorylates specific protein targets, including ion channels; after translocating to the nucleus, Hog1 phosphorylates several transcription factors and chromatin-modifying enzymes. Hog1 also appears also to promote transcriptional elongation. Together, the newly synthesized and modified proteins support survival in high-osmotic-strength media. See N. Dard and M. Peter, 2006, BioEssays28:146, and R. Chen and J. Thorner, 2007, Biochim. Biophys. Acta1773:1311.
In yeast mating pathways, Gβγ functions by triggering a kinase cascade that is analogous to the one downstream from Ras; each protein in the cascade has a yeast-specific name, but shares sequences with, and is analogous in structure and function to, the corresponding mammalian protein shown in Figure 16-24. The components of this cascade were uncovered mainly through analyses of mutants that possess functional a and α receptors and G proteins, but are sterile (Ste) or defective in mating responses. The physical interactions between the components of the cascade were assessed through immunoprecipitation experiments with extracts of yeast cells and other types of studies. Based on these studies, scientists have proposed the kinase cascade shown in Figure 16-27a. Free Gβγ, which is tethered to the membrane via the lipid bound to the γ subunit, binds the Ste5 protein, thus recruiting it and its bound kinases to the plasma membrane. Ste5, which has no obvious catalytic function, acts as a scaffold for assembling other components in the cascade (Ste11, Ste7, and Fus3). Gβγ also activates cdc24, a GEF for the Ras-like protein Cdc42; GTP·Cdc42 in turn activates the Ste20 protein kinase. Ste20 phosphorylates and activates Ste11, a serine/threonine kinase analogous to Raf and other mammalian MEK kinase (MEKK) proteins. Activated Ste11 then phosphorylates Ste7, a dual-specificity MEK, which then phosphorylates and activates Fus3, a serine/threonine kinase equivalent to MAP kinase. After translocation to the nucleus, Fus3 phosphorylates two proteins, Dig1 and Dig2, relieving their inhibition of the Ste12 transcription factor. Activated Ste12, in turn, induces expression of proteins involved in mating-specific cellular responses. Fus3 also affects gene expression by phosphorylating other proteins.