Hedgehog Signaling Relieves Repression of Target Genes
The Hedgehog (Hh) signaling pathway is similar to the Wnt pathway in that two membrane proteins, one with seven membrane-spanning segments, are required to receive and transduce a signal. The Hh pathway also involves the disassembly of an intracellular complex containing a transcription factor. Hh signaling differs from Wnt signaling in that its two membrane receptors move between the plasma membrane and intracellular vesicles, and that in mammals, Hh signaling is restricted to the primary cilium that protrudes from the cell surface. But, like Wnt proteins, Hh proteins can act as morphogens and signal nearby cells. Hh signaling plays essential roles in the development of nearly every organ system in vertebrates, from determining the fates of different segments of the nervous system to regulating lung morphogenesis and hair follicle formation. One of the three mammalian Hh proteins, Sonic Hedgehog (Shh), is essential for normal patterning of the limbs; abnormal expression of Shh in an anterior region of the developing limb, in addition to its normal expression in the posterior domain, leads to polydactyly (extra digits).
Page 755
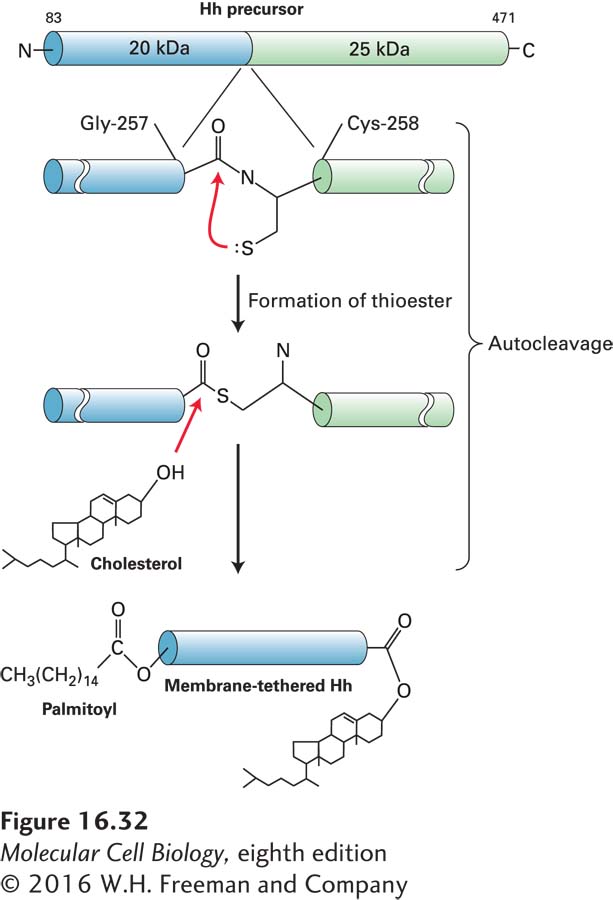
Processing of Hh Precursor Protein Hedgehog proteins are formed from a precursor protein with autoproteolytic activity that enables the protein to cut itself in half while still in the endoplasmic reticulum. The cleavage produces an N-terminal fragment, which is subsequently secreted to signal other cells, and a C-terminal fragment, which is degraded. As shown in Figure 16-32, cleavage of the precursor is accompanied by covalent addition of the lipid cholesterol to the new carboxyl terminus of the N-terminal fragment. A second modification to Hedgehog, the addition of a palmitoyl group to the N-terminus, makes the protein even more hydrophobic.
FIGURE 16-32Processing of Hedgehog (Hh) precursor protein. Cells synthesize a 45-kDa Hh precursor, which in the endoplasmic reticulum undergoes a nucleophilic attack by the thiol side chain of cysteine 258 (Cys-258) on the carbonyl carbon of the adjacent residue glycine 257 (Gly-257), forming a high-energy thioester intermediate. Enzyme activity in the C-terminal domain then catalyzes the formation of an ester bond between the hydroxyl group of cholesterol and glycine 257, cleaving the precursor into two fragments. The N-terminal signaling fragment (blue) retains the cholesterol group and is also modified by the addition of a palmitoyl group to the N-terminus. The two hydrophobic anchors may tether the secreted, processed Hh protein to the plasma membrane. See P. Thérond, 2012, Curr. Opin. Cell Biol.24:173.
Hh proteins can travel relatively long distances—up to 300 µm in the developing vertebrate limb—but how an Hh protein with two attached lipids can spread in the hydrophilic environment of the extracellular space is not well understood. Both Hh and Wnt are found anchored to the phospholipid monolayers of extracellular lipoprotein particles (see Figure 14-27 for the structure of a typical lipoprotein) via their attached lipid or cholesterol groups. In many cases, the majority of the Hh proteins produced by a cell remain bound to its plasma membrane; in such cases, Hh signals mainly by cell-cell contact. Extracellular aggregates of Hh proteins, stabilized by hydrophobic interactions between their lipid groups, have also been observed experimentally. Regardless of the detailed mechanism involved, the two attached hydrophobic groups limit the diffusion of Hh and thus its range of action in tissues. As with Wnt proteins, spatial restriction plays a crucial role in constraining the effects of Hh proteins.
Hedgehog Signaling Pathway in Drosophila Genetic studies in Drosophila indicated that two membrane proteins, Smoothened (Smo) and Patched (Ptc), are required to receive a Hedgehog signal and transduce it to the cell nucleus. Smoothened has seven membrane-spanning α helices and is related in sequence to the Wnt receptor Fz. Patched is predicted to contain twelve transmembrane α helices and is most similar structurally to the Niemann-Pick C1 (NPC1) protein, a member of the ABC superfamily of membrane transport proteins (see Table 11-3).
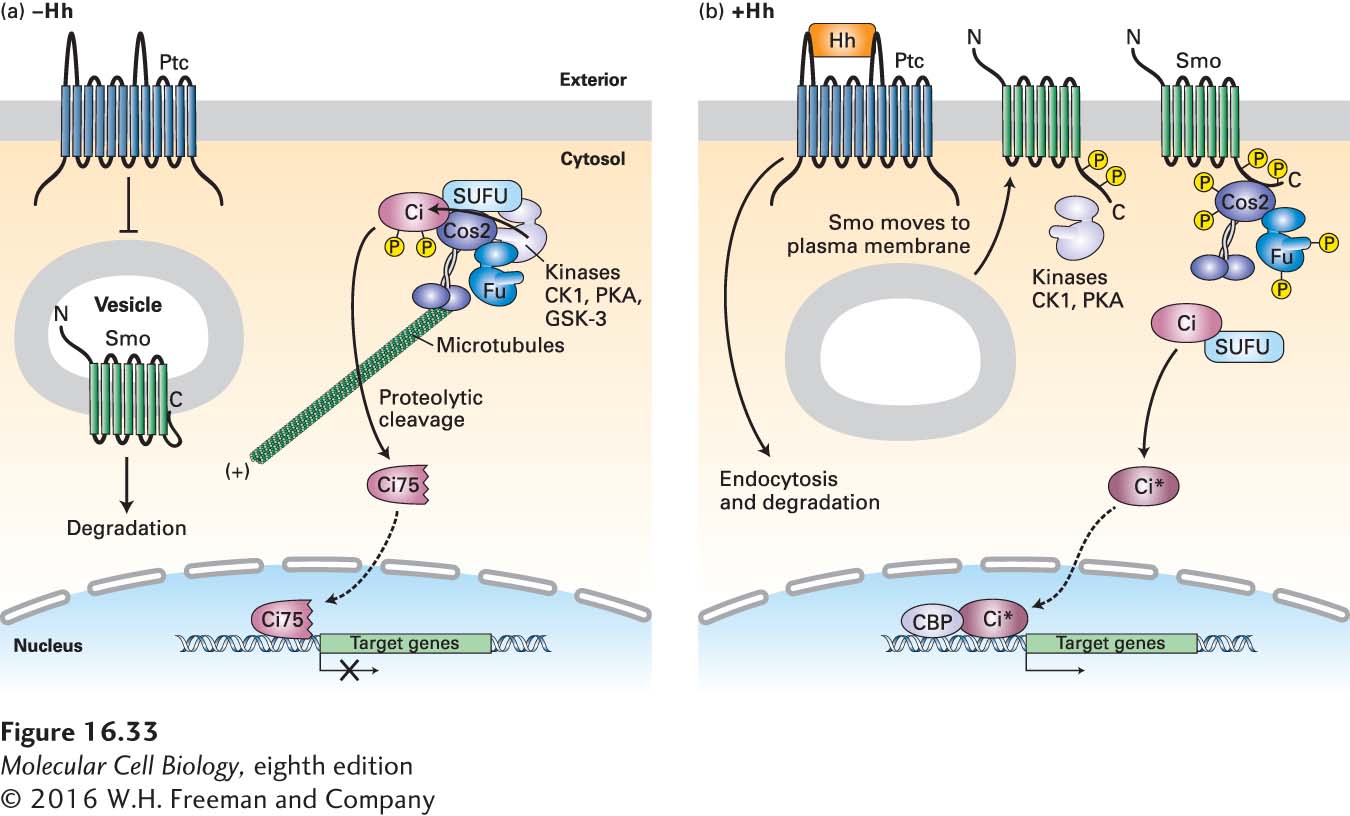
Figure 16-33 depicts the current model of the Hedgehog pathway in Drosophila. Evidence supporting this model initially came from the study of fly embryos with loss-of-function mutations in the hedgehog (Hh) or smoothened (Smo) genes. Both types of mutant embryos have very similar developmental phenotypes; the name hedgehog came from the appearance of Hh mutant embryos, which were covered by an array of disorganized hairlike bristles that resembled hedgehog spines. Moreover, both the Hh and Smo genes are required to activate transcription of the same target genes (e.g., patched and wingless) during embryonic development. In contrast, loss-of-function mutations in the patched (Ptc) gene produce a quite different phenotype, one similar to the effect of flooding the embryo with Hedgehog protein. These findings suggested that, in the absence of Hh, Ptc represses target genes by inhibiting a signaling pathway needed for gene activation. The additional observation that Smo is required for the transcription of Hh target genes in mutants lacking patched function places Smo downstream of Ptc in the Hh pathway. Together with other experiments showing that Hh binds directly to Ptc, the evidence indicates that Hh binding to Ptc prevents Ptc from blocking Smo action, thus activating the transcription of target genes.
FIGURE 16-33Hedgehog signaling in Drosophila. (a) In the absence of Hedgehog (Hh), Patched (Ptc) protein inhibits Smoothened (Smo), which is present largely in the membranes of internal vesicles, and enhances its degradation. A complex containing the kinase Fused (Fu); other kinases including protein kinase A (PKA), glycogen synthase kinase 3β (GSK3β), and casein kinase 1 (CK1); the kinesin-related motor protein Costal-2 (Cos2); and Cubitis interruptus (Ci), a zinc-finger transcription factor, binds to microtubules. In this complex, Ci becomes phosphorylated in a series of steps catalyzed by PKA, GSK3β, and CK1. The phosphorylated Ci is then proteolytically cleaved by the ubiquitin-proteasome pathway, generating the N-terminal fragment Ci75, which is transported into the nucleus and functions as a transcriptional repressor of Hh target genes. (b) Hh binds to Ptc, causing Ptc to be endocytosed from the cell surface and degraded, thereby relieving the inhibition of Smo. Smo then moves to the plasma membrane, is phosphorylated by PKA, CK1, and other kinases, binds Cos2, and is stabilized from degradation. Both Fu and Cos2 become extensively phosphorylated, and most importantly, the Fu-Cos2-Ci complex becomes dissociated. This leads to the stabilization of the full-length Ci, which moves into the nucleus, displaces the repressor Ci75 from the promoter of target genes, recruits the CREB-binding activator protein (CBP), and induces expression of target genes. The exact membrane compartments in which Ptc and Smo respond to Hh and function are unknown. See S. Goetz and K. Anderson, 2010, Nat. Rev. Genet.11:331.
Page 756
Subsequent biochemical and cell biological studies showed that, in the absence of Hh, Ptc is enriched in the plasma membrane, but Smo is found in membranes of internal vesicles. Furthermore, the long cytosolic C-terminal segment of Smo is folded in such a way that it cannot bind downstream signaling proteins (Figure 16-33a). How Ptc inhibits Smo function and enhances its degradation is not clear; one theory is that Ptc transports a small molecule into the cell that inhibits Smo function.
A large cytosolic protein complex in the Hh pathway consists of several proteins including Fused (Fu), a serine-threonine kinase; protein kinases PKA, GSK3β, and CK1, which we have encountered previously in other signaling pathways; Costal-2 (Cos2), a microtubule-associated kinesin-like protein; and Cubitis interruptus (Ci), a zinc finger–containing transcription factor. The complex is bound to, and may move along, microtubules in the cytosol. Importantly, phosphorylation of Ci by at least three kinases in the complex causes binding of a component of a ubiquitin ligase complex, which in turn directs ubiquitinylation of Ci and its targeting to proteasomes. There Ci undergoes proteolytic cleavage; the resulting Ci fragment, designated Ci75, translocates to the nucleus and represses expression of Hh target genes.
Following binding of Hh to the receptor Ptc, the Hh-Ptc complex, like other receptor-hormone complexes, is endocytosed from the cell surface into internal vesicles and is eventually degraded; the binding of Hh to Ptc also inhibits the ability of Ptc to inhibit Smo (see Figure 16-33a). Simultaneously, Smo moves from internal vesicles to the plasma membrane; the C-terminal segment of Smo becomes phosphorylated and adopts an “open” conformation. This change triggers several cellular responses, including an increase in phosphorylation of Fu and Cos2. Importantly, the complex of Fu, Cos2, and Ci dissociates from microtubules, and Cos2 becomes associated with the phosphorylated C-terminal tail of Smo. The resulting disruption of the Fus/Cos2/Ci complex causes a reduction in both phosphorylation and cleavage of Ci. As a result, full-length Ci is released and translocates to the nucleus, where it binds to the transcriptional co-activator CREB-binding protein (CBP), promoting the expression rather than repression of Hh target genes.
Page 757
Regulation of Hh Signaling Feedback control of the Hh pathway is important because unrestrained Hh signaling can cause cancerous overgrowth or formation of the wrong cell types. In Drosophila, one of the genes induced by the Hh signal is patched. The subsequent increase in expression of Patched antagonizes the Hh signal in large measure by reducing the pool of active Smoothened protein. Thus the system is buffered: if during development too much Hh signal is made, a consequent increase in Ptc will compensate; if too little Hh signal is made, the amount of Ptc is decreased.