Actin Filament Treadmilling Is Accelerated by Profilin and Cofilin
Measurements of the rate of actin treadmilling in vivo show that it can be several times higher than can be achieved with pure actin in vitro under physiological conditions. Consistent with the treadmilling model, growth of actin filaments in vivo occurs only at the (+) end. How is enhanced treadmilling achieved, and how does the cell recharge the ADP-actin dissociating from the (−) end to ATP-actin for assembly at the (+) end? Two different actin-binding proteins make important contributions to these processes.
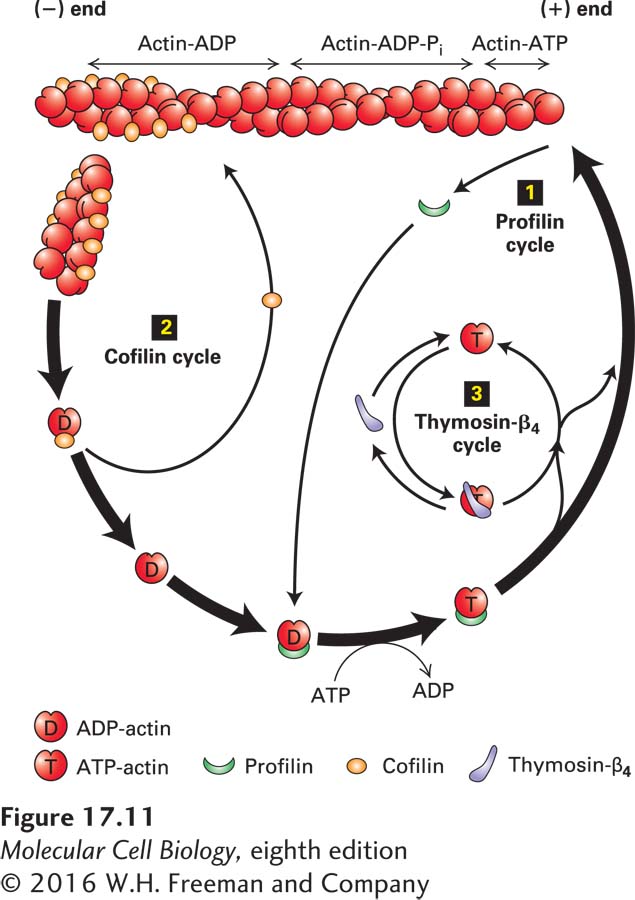
The first of these proteins is profilin, a small protein that binds G-actin on the side opposite the nucleotide-binding cleft. When profilin binds ADP-actin, it opens the cleft and greatly enhances the loss of ADP, which is replaced by the more abundant cellular ATP, yielding a profilin–ATP-actin complex. This complex cannot bind to the (−) end because profilin blocks the sites on G-actin for (−) end assembly. However, the profilin–ATP-actin complex can bind efficiently to the (+) end, and profilin dissociates after the new actin subunit is bound (Figure 17-11). This function of profilin on its own does not enhance the treadmilling rate, but it does provide a supply of ATP-actin from released ADP-actin; as a consequence, essentially all the free G-actin in a cell has bound ATP.
FIGURE 17-11Regulation of filament turnover by actin-binding proteins. Actin-binding proteins regulate the rate of assembly and disassembly of actin filaments as well as the availability of G-actin for polymerization. In the profilin cycle 1, profilin binds ADP–G-actin and catalyzes the exchange of ADP for ATP. The ATP–G-actin–profilin complex can deliver actin to the (+) end of a filament with dissociation and recycling of profilin. In the cofilin cycle 2, cofilin binds preferentially to filaments containing ADP-actin, inducing them to fragment and thus enhancing depolymerization by making more filament ends. In the thymosin-β4 cycle 3, ATP–G-actin made available by the profilin cycle is bound by thymosin-β4, which sequesters it from polymerization. As the free G-actin concentration is lowered by polymerization, G-actin–thymosin-β4 dissociates to make free G-actin available for association with profilin and further polymerization.
Profilin has another important property: it can bind other proteins with sequences rich in proline residues at the same time it is binding actin. We will see later how this property functions in actin filament assembly.
Cofilin is another small protein involved in actin treadmilling, but it binds specifically to F-actin in which the subunits contain ADP, which are the older subunits in the filament toward the (−) end (see Figure 17-10a). Cofilin binds by bridging two actin monomers and inducing a small change in the twist of the filament. This small twist destabilizes the filament, breaking it into short pieces. By breaking the filament in this way, cofilin generates many more free (−) ends and therefore greatly enhances the net disassembly of the filament (see Figure 17-11). The released ADP-actin subunits are then recharged by profilin and added to the (+) end as described above. In this way, profilin and cofilin can enhance treadmilling in vitro more than tenfold, up to levels seen in vivo. As might be anticipated, the cell uses signal transduction pathways to regulate both profilin and cofilin, and thereby the turnover of actin filaments.